Освіта жовчі. Фізіологія желчеобразования. Освіта жовчі
Освіта жовчі відбувається в печінці в результаті активного і пасивного транспорту речовин (вода, глюкоза, креатинін, електроліти, гормони, вітаміни та ін.), Що циркулюють в крові, через клітини і міжклітинні контакти, а також активної секреції компонентів жовчі (жовчні кислоти) гепатоцитами і зворотного всмоктування води і деяких речовин з дрібних жовчних проток і ЖП. Фізіологічне значення цього процесу різноманітне. Жовч насамперед розглядають як травний секрет, оскільки жовчні кислоти (переважно їх органічні аніони) відіграють ключову роль в абсорбції жирів. Жовч емульгує жири, збільшуючи поверхню, на якій здійснюється їх гідроліз під впливом ліпази. Під дією жовчі відбувається розчинення продуктів гідролізу жирів. Вона сприяє їх всмоктуванню і ресинтезу тригліцеридів в ентероцитах. Жовч підвищує активність ферментів ПЖ і кишечника (особливо ліпази), підсилює гідроліз і всмоктування білків і вуглеводів.
При порушенні перетравлення жирів погано перетравлюються і інші харчові речовини, тому що жир обволікає частинки їжі і перешкоджає дії на них ферментів. У подібних умовах діяльність кишкових бактерій призводить до посилення процесів гниття, бродіння і утворення газу.
Жовчі притаманне і регуляторну дію - стимуляція жовчоутворення, жовчовиділення, моторної і секреторної діяльності тонкої кишки, а також проліферації і злущування ентероцитів. Жовч гальмує процес шлункового травлення шляхом нейтралізації кислоти, що надійшла в ДПК, і інактивації пепсину, готуючи до травленню в кишечнику. Вона також надає регуляторний вплив на евакуаторну діяльність гастродуоденальної комплексу. Важлива роль жовчі у всмоктуванні з просвіту кишечника жиророзчинних вітамінів (А, Д, Е і К), холестерину, солей кальцію.
Крім того, утворення і виділення жовчі розглядають як своєрідний шлях виведення з організму деяких молекул і іонів, які не можуть бути виділені через нирки. Серед них найбільш значущими є холестерин (виводиться у вигляді вільного холестерину, його ефірів і жовчних кислот), білірубін, а також молекули міді і заліза. Тому жовч розглядають і як екскреторну рідина.
Жовч складається з 80% води і 20% розчинених в ній речовин. Останні включають жовчні кислоти і їх солі (близько 65%), біліарний фосфоліпіди (близько 20%, в основному за рахунок лецитину), білки (близько 5%), холестерин (4%), кон'югований білірубін (0,3%), ферменти , імуноглобуліни, а також численні екзогенні та ендогенні речовини, що виділяються з жовчю (рослинні стироли, вітаміни, гормони, лікарські препарати, токсини, іони металів - міді, заліза, калію, натрію, кальцію, цинку, магнію, ртуті та ін.). В середньому на добу секретується 600-700 мл жовчі (від 250 до 1500 мл, приблизно 10,5 мл на 1 кг маси тіла). При цьому близько 500 мл / сут загального обсягу забезпечується секрецією гепатоцитів і близько 150 мл / сут секретують клітини жовчних проток.
Освіта жовчі (Холерез) йде безперервно, а надходження жовчі в ДПК (Холекінез) відбувається періодично. Натщесерце жовч в кишку надходить відповідно до голодної періодичної діяльністю. У періоди спокою вона направляється в ЖП, де концентрується, трохи змінює свій склад і депонується. Крім води і солей, абсорбуються холестерин і вільні жирні кислоти. У зв'язку з цим розрізняють печінкову і міхурово жовч.
Жовч має невелику ферментативної активністю; рН печінкової жовчі становить 7,3-8,0. На відміну від кишкового вмісту вона практично не містить бактерій. До чинників, які забезпечують стерильність жовчі, відносять наявність жовчних кислот (бактеріостатичну дію), багатий зміст імуноглобулінів, секрецію слизу, відносну бідність жовчі енергетичними субстратами для бактерій.
Жовч є мицеллярний розчин. Холестерин, практично не розчинний у воді, транспортується в розчиненому стані в жовчі завдяки її мицеллярная будовою. Цей процес називають колоїдним розчиненням - Солюбілізація.
Жовчні кислоти є поверхнево-активними, амфіпатіческімі (одночасно гідрофобні і гідрофільні) молекулами, здатними до самоагрегаціі. При цьому завдяки дуже вузькій концентраційної нормі, званої критичної концентрацією міцеллізаціі, утворюються прості міцели. Прості міцели мають виражену здатність до розчинення ліпідів, формуючи змішані міцели.
Вважають, що змішані міцели мають циліндричне будова: циліндричний стовбур заповнений полярними ліпідами, а молекули жовчних кислот розташовуються між полярними кінцями ліпідних молекул з гідрофільними сторонами, зверненими до водного оточення, що й обумовлює їх гідрофільність (водорастворимость).
Змішані міцели містять обов'язкові компоненти - жовчні кислоти, розташовані зовні, фосфоліпіди (в основному фосфатидилхолін - лецитин) і холестерин, розташовані всередині міцели.
Колір жовчі - жовтувато-коричневий через присутність білірубіну, найбільша частина якого представлена у вигляді диглюкороніду білірубіну, менша - у вигляді моноглюкороніду білірубіну. Пов'язаний білірубін не входить до складу мицелл.
Оскільки міжклітинні з'єднання біліарного дерева проникні для води, міхурна і печінкова жовч ізотонічни.
жовчоутворення
билиарная секреція
Жовчні кислоти є основним компонентом біліарної секреції, вони утворюються тільки в печінці. Первинні жовчні кислоти - трігідроксіхолевая (холіевая) і дігідроксіхолевая (хенодезоксихолева) синтезуються в гепатоцитах з холестерину. Вторинні жовчні кислоти (Дезоксихолева і в малих кількостях - літохолевая) утворюються в товстій кишці з первинних в результаті бактеріальної модифікації ядерних гідроксильних груп (7а-дегідроксілірованіе під контролем анаеробних бактерій). Третинні жовчні кислоти (В основному урсодезоксихолевая) утворюються в печінці шляхом ізомеризації вторинних жовчних кислот.
Синтез жовчних кислот з холестерину регулюється механізмом негативного зворотного зв'язку: знижений повернення жовчних кислот в гепатоцити з портальної кров'ю призводить до збільшення їх біосинтезу; будь-яке збільшення синтезу жовчних кислот супроводжується відповідним підвищенням освіти холестерину.
Жовчні кислоти кон'югують в печінці з амінокислотами гліцином (близько 80%) або таурином (близько 20%). При вживанні їжі, багатої вуглеводами, в жовчі зростає кількість глікохолевой кислот, при високобілковою дієті - таурохолевой. Зв'язування з амінокислотами запобігає всмоктуванню жовчних кислот в жовчних шляхах і початкових відділах тонкої кишки (тільки в термінальному відділі клубової кишки і в товстій кишці). Під дією бактерій може бути гідроліз солей жовчних кислот з утворенням жовчних кислот, гліцину або таурину.
Після біосинтезу жовчних кислот, як було зазначено вище, відбувається зв'язування їх карбоксильної групи з аминогруппой гліцину або таурину. З хімічної точки зору таке зв'язування конвертує слабку кислоту в сильнішу. Пов'язані жовчні кислоти найбільш розчинні при низькому рН і більш стійкі до осадження іонами кальцію (Ca 2+), ніж незв'язані жовчні кислоти. З біологічної ж точки зору процес зв'язування робить жовчні кислоти нездатними до проникнення через клітинні мембрани. Тому вони абсорбуються в жовчних протоках або тонкій кишці або в присутності молекул-носіїв, або якщо жовчні кислоти піддаються бактеріального розщеплення. Єдиним винятком з цього правила є гліціндігідроксісвязанние жовчні кислоти, оскільки вони можуть абсорбуватися пасивно, якщо отримують іон водню (Н +).
Велика частина жовчних кислот (близько 85-90%), що надійшли з струмом жовчі в тонку кишку, всмоктується в кров. При цьому більшість пов'язаних жовчних кислот, які секретуються в тонку кишку, абсорбується інтактними. Менша частина кислот не абсорбується інтактними, оскільки піддається бактеріальному розщепленню в дистальних відділах тонкої кишки. Вони всмоктуються пасивно і, повертаючись в печінку, знову зв'язуються і секретуються в жовч. Решта 10-15% жовчних кислот виводяться з організму в основному з калом. Ця втрата жовчних кислот компенсується їх синтезом в гепатоцитах.
Цей процес кишкового розщеплення і повторного всмоктування в печінці є нормальною складовою частиною метаболізму жовчних кислот. Невелика частина жовчних кислот, які не зазнала абсорбції, надходить в товсту кишку. Тут закінчується процес розщеплення. Крім того, в товстій кишці відбувається утворення вторинних жовчних кислот під дією анаеробних бактерій (див. Вище).
Дезоксихолева і літохолевая кислоти частково всмоктуються в товстій кишці і знову надходять в печінку. Після повернення метаболізм цих жовчних кислот різний. Дезоксихолева кислота зв'язується з гліцином або таурином і циркулює разом з первинними жовчними кислотами. Слід зазначити, що у літніх людей дезоксихолева кислота є основною жовчної кислотою в складі жовчі. Литохолевая кислота не тільки зв'язується з гліцином або таурином, а й додатково сульфатується в положенні С-3. Таке «подвійне» зв'язування знижує можливість всмоктування в кишечнику, в результаті чого літохолевая кислота швидко втрачається зі складу циркулюючих жовчних кислот - в жовчі її зміст рідко перевищує 5%. Більшу частину (95%) жовчних кислот, що входять до складу жовчі дорослих людей, складають холевая, хенодезоксихолева і дезоксихолева кислоти.
Процес зв'язування жовчних кислот в гепатоцитах вельми ефективний, тому біліарние жовчні кислоти повністю представлені в кон'югованої формі. Подальше бактеріальне розщеплення і дегідроксілірованіе призводять до того, що в калі жовчні кислоти знаходяться в незв'язаному стані.
Описаний вище процес дегідроксілірованія первинних жовчних кислот знижує їх здатність до розчинення у воді. Освіта вторинних жовчних кислот у людини практично не має фізіологічного значення на відміну від тварин. Надмірна ж абсорбція дезоксихолевої кислоти в товстій кишці підвищує ризик утворення холестеринових жовчних каменів. Літохолевую кислоту вважають гепатотоксичності. В експериментах на тваринах показано, що накопичення литохолевой кислоти в печінці призводить до її поразки. Разом з тим поки не доведено, що підвищення абсорбції цієї кислоти у людини веде до порушення роботи печінки.
Урсодезоксихолева кислота, так само як і дезоксихолева, зв'язується в печінці і циркулює разом з первинними жовчними кислотами. Однак шлях метаболізму цієї кислоти дуже короткий і зміст пов'язаної урсодезоксихолевої кислоти в жовчі ніколи не перевищує 5% від загальної кількості жовчних кислот. Вважають, що освіта урсодезоксихолевої кислоти не має важливого фізіологічного значення.
Жовчні кислоти є потужними розчинниками жирів, тому вони цитотоксичних в концентраціях, що досягають критичної концентрації міцеллізаціі. При цьому пов'язані хенодезоксихолева і дезоксихолева кислоти, присутні в жовчі, більш токсичні, ніж пов'язані холевая і урсодезоксихолевая. Хоча жовчні кислоти демонстративно цитотоксичних in vitro, епітелій жовчних проток і тонкої кишки ніколи не пошкоджується в результаті їх високої концентрації. Це пов'язують, по-перше, з присутністю інших ліпідів (фосфоліпіди в жовчі і жирні кислоти в кишечнику), що знижують монометріческую концентрацію жовчних кислот, і, по-друге, з наявністю гликолипидов і холестерину в апікальних мембранах епітеліальних клітин, підвищують резистентність епітеліоцитів до шкідлива впливу аніонів жовчних кислот.
Холевая, хенодезоксихолева і дезоксихолева кислоти всмоктуються і піддаються ентерогепатичній циркуляції до 6-10 раз на добу. Литохолевая кислота всмоктується погано і її кількість в жовчі мало. Пул жовчних кислот становить в нормі близько 2,5 г, а добова продукція первинних жовчних кислот - холевой і хенодезоксихолевої - в середньому близько 330 і 280 мг відповідно.
регуляція жовчоутворення
Як відомо, желчеобразование відбувається безперервно, однак інтенсивність цього процесу змінюється. Підсилюють желчеобразование прийом їжі і прийнята їжа. Рефлекторний вплив на холерез відбувається при подразненні рецепторів шлунково-кишкового тракту (ШКТ), внутрішніх органів і умовно-рефлекторному впливі.
Вегетативна регуляція забезпечується парасимпатическими холинергическими (підсилюють жовчоутворення) і симпатичними адренергічними нервовими волокнами (зменшують жовчоутворення).
Гуморальна регуляція здійснюється самою жовчю завдяки наявності ентерогепатичній циркуляції жовчних кислот і механізму зворотного негативного зв'язку (див. Вище). Секретин стимулює секрецію жовчі, виділення в її складі води і електролітів. Більш слабке стимулюючий вплив надають глюкагон, гастрин і холецистокінін.
билиарная екскреція
Билиарную екскрецію розглядають як своєрідний шлях видалення з організму молекул і іонів, які не можуть бути виділені через нирки. Найбільш важливими серед них вважають холестерин (як такої і у вигляді жовчних кислот) і білірубін, а також іони міді, заліза та ін.
Основні компоненти біліарної екскреції
холестерин, Практично не розчинний у воді, транспортується в складі змішаних міцел, утворених, як було зазначено вище, жовчними кислотами, фосфоліпідами і власне холестерином.
Молекули фосфоліпідів виконують дві важливі функції. По-перше, вони істотно збільшують міцелярно розчинність холестерину, оскільки змішані міцели, що містять фосфоліпіди, солюбілізіруют набагато більше холестерину, ніж прості міцели, що містять лише молекули жовчних кислот. По-друге, наявність фосфоліпідів в жовчі зменшує критичну концентрацію міцеллізаціі і монометріческую концентрацію жовчних кислот. В результаті цього знижується поверхнева активність і цитотоксичність печінкової жовчі.
З потоком жовчі змішані міцели надходять в тонку кишку, де подальше перетворення складових їх компонентів різна. Жовчні кислоти розчиняють ліпіди, забезпечуючи їх абсорбцію, і самі абсорбуються в більш дистальних відділах кишечника. Фосфоліпіди жовчі, нерозчинні у воді, гідролізуються в кишечнику і не залучаються до ентерогепатичну циркуляцію. Жовчні кислоти регулюють їх виведення і стимулюють синтез.
Оскільки абсорбується приблизно 1/3 холестерину, 2/3 його виділяється. У дорослої людини баланс холестерину забезпечується його виділенням як такого (близько 600 мг / добу) або у вигляді жовчних кислот (близько 400 мг / добу). При цьому ентерогепатичну циркуляцію жовчних кислот розглядають як уповільнений шлях виведення холестерину.
Слід зазначити, що в порівнянні з тваринами у людини відзначається більш висока пропорція холестерину в жовчі. Вони вважають його дуже результатом дефективної конверсії холестерину в жовчні кислоти, а також і відносно низькою швидкості секреції жовчних кислот. Тому жовч виявляється перенасиченої холестерином приблизно у 25% літніх людей, а у 10-15% населення утворюються холестеринові камені (холелітіаз).
Як було зазначено вище, синтез жовчних кислот з холестерину регулюється механізмом зворотного негативного зв'язку.
Жовч є основним шляхом виведення білірубіну з організму. білірубін - кінцевий продукт розпаду гема, за своєю хімічною структурою являє тетрапіррол. Найбільша кількість білірубіну (80-85%) відбувається з гемоглобіну, що розпався старіючих еритроцитів і передчасно руйнуються новостворених еритроцитів в кістковому мозку або циркуляторном руслі (так званий неефективний еритропоез). Інша частина білірубіну утворюється в результаті руйнування інших гемсодержащих білків (наприклад, цитохрому Р-450 і ін.) В печінці і набагато менше - у внепеченочной тканини. Слід зазначити, що катаболізм гемоглобіну еритроцитарної природи в білірубін в першу чергу відбувається в макрофагах селезінки, печінки і кісткового мозку.
Джерела утворення білірубіну (в процентному співвідношенні)

Як відомо, некон'югований білірубін є гідрофобним (нерозчинним у воді) і потенційно токсичною речовиною, яке циркулює в плазмі в тісно пов'язаному з альбуміном стані і не може бути екскретуватися з сечею. Можливості організму по виведенню білірубіну пов'язані з видаленням останнього з плазми крові печінкової клітиною, наступним кон'югірованію з глюкуроновою кислотою і виділенням в жовч вже водорозчинного з'єднання (пов'язаного або прямого білірубіну). Процес кон'югації протікає за допомогою мікросомального ферменту урідіндіфосфатглюкуронілтрансферази. Кон'югація білірубіну з глюкуроновою кислотою має найбільш важливе фізіологічне значення, проте в невеликих кількостях відбувається кон'югування білірубіну з сульфатами, глюкозою і ксилозой.
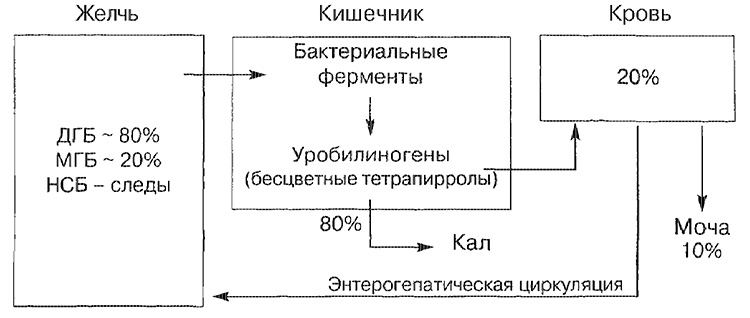
Основні шляхи транспорту білірубіну
НСБ - непов'язаний білірубін; МГБ - моноглюкуронід білірубіну; ДГБ - диглюкуронід білірубіну.
Виділення кон'югованого білірубіну в жовчні канальці відбувається за участю сімейства АТФ-залежних мультіспеціфічних транспортних білків для органічних аніонів.
Найбільша частина білірубіну жовчі (близько 80%) представлена в формі диглюкороніду білірубіну, менша - у вигляді моноглюкороніду і лише незначна його кількість представлено незв'язаної формою. Білірубін, що надійшов в кишечник, розщеплюючи в термінальному відділі тонкої кишки і в товстій кишці бактеріальними ферментами (бета-глюкуронідазу), конвертується в безбарвні тетрапірроли (уробіліноген). Близько 20% утворилися уробіліногенов резорбируется і обов'язково виводиться з сечею, а також з жовчю (ентерогепатичній коло циркуляції).
Більшість авторів сходяться на думці, що нормальна концентрація загального білірубіну сироватки крові, певна діазореакція по Van Der Bergh, зазвичай не перевищує 1 мг% (0,3-1 мг%, або 5-17 мкмоль / л). Лише менше 5% білірубіну представлено у вигляді пов'язаної форми. Підвищення рівня білірубіну в крові (гіпербілірубінемія) і накопичення його в тканинах призводять до появи жовтяниці, яка, як правило, стає помітною при значеннях, що перевищують 2,5-3 мг%.
Як було зазначено вище, білірубін циркулює в крові у зв'язаному з білком стані і насилу проникає в тканинні рідини з низьким вмістом білка. У зв'язку з цим ексудати мають більш жовте забарвлення, ніж транссудату. Білірубін добре зв'язується з еластичної тканиною, що пояснює рання жовте забарвлення склер, шкіри та стінок судин при гіпербілірубінемії. Тривалий холестаз призводить до появи зеленуватого відтінку шкіри, що пояснюють відкладенням биливердина.
Доречно буде підкреслити, що тиск в жовчних протоках, на тлі якого відбувається секреція жовчі, в нормі становить 150-200 мм вод. ст. Його збільшення до 350 мм вод. ст. пригнічує секрецію жовчі, приводячи до розвитку жовтяниці. У разі повного припинення секреції білірубіну і жовчних кислот жовч знебарвлюється (так звана біла жовч).
Крім того, жовч є шляхом виведення з організму рослинних жирів, ліпофільних лікарських препаратів та їх метаболітів, різних ксенобіотиків, які присутні в рослинах, ліпофільних метаболітів, жиророзчинних вітамінів і стероїдних гормонів.
Баланс заліза та міді в організмі також підтримується завдяки біліарної екскреції цих металів. Обидва катіона виділяються в жовч за допомогою АТФ-стимульованого каналікулярного насоса. З жовчю відбувається виділення і інших металів.
желчевиведеніе
Струм жовчі в жовчовивідної системі обумовлений різницею тиску в різних її відділах і ДПК, тонусом сфінктерів, скороченням гладких волокон ЖП і проток (перші окремі м'язові волокна з'являються у внутрішньопечінковий жовчних протоках, розташованих в безпосередній близькості від воріт печінки). Діяльність всієї системи в нормі добре скоординована і знаходиться під контролем нервових і гуморальних механізмів.
В ЖП відбувається депонування і підвищення концентрації жовчі з 10-кратним збільшенням концентрації ліпідів, солей жовчних кислот, жовчних пігментів і ін. В потрібний момент здійснюється її викид в ОЖП і далі в ДПК. Пасаж жовчі координує сфінктерного апарату. Поза процесом травлення в ДПК сфінктер ампули закритий. В цей час ЖП і протоки міхура розслаблені, що дозволяє жовчі, що виробляється печінкою, надходити в просвіт ЖП. Слизова оболонка ЖП всмоктує воду, іони. При цьому жовч стає більш концентрованою. Продукція слизу дозволяє жовчі перебувати в колоїдному стані.
Тиск в просвіті ЖМ в спокої набагато менше, ніж в жовчних протоках, і становить 60-185 мм вод. ст. Різниця в тиску є фізіологічною основою надходження жовчі в ЖП при закритому сфінктера Одді. В процесі травлення за рахунок скорочення ЖП тиск зростає до 150-260 мм вод. ст, забезпечуючи надходження жовчі в ДПК через розслаблений сфінктер ампули. Коли жовч починає надходити в ДПК, тиск в протоках поступово знижується (при кожному прийомі їжі ЖП скорочується 1-2 рази).
Період первинної реакції жовчовидільної системи, обумовлений видом, запахом їжі і її прийомом, триває приблизно 7-10 хв. Потім настає евакуаторної період (основний або період спорожнення ЖП), протягом якого на тлі чергування скорочень і розслаблень ЖП в ДПК виливається спочатку міхурна, а потім і печінкова жовч.
Рефлекторна стимуляція холекінез (умовно і безумовно-рефлекторна) відбувається завдяки рецепторам, що знаходяться в порожнині рота, шлунку і ДПК, за допомогою блукаючих нервів. Гуморальна регуляція в основному здійснюється під впливом холецистокініну, який має стимулюючий вплив на ЖП, викликаючи його скорочення. Крім того, слабкі скорочення ЖП відбуваються під впливом гастрину, секретину і бомбезіна. Навпаки, гальмують скорочення ЖП глюкагон, кальцитонін, антіхолецістокінін, вазоінтестінальний пептид і панкреатичний поліпептид.
Потужними стимуляторами жовчовиділення є яєчні жовтки, жири, молоко і м'ясо.
Порушення діяльності сфінктерного апарату і моторики ЖП можуть призводити до зміни спрямованості і швидкості струму жовчі, дискінезії, рефлюксу секрету ПЖ і вмісту ДПК в ОЖП, а жовчі - в панкреатичний протік, що є основою для розвитку ряду патологічних станів і захворювань.
Солі жовчних кислот, кон'югований білірубін, холестерин, фосфоліпіди, білки, електроліти та вода секретируются гепатоцитами в жовчні канальці. Апарат секреції жовчі включає в себе транспортні білки канальцевої мембрани, внутрішньоклітинні органелиі структури цитоскелета. щільні контактиміж гепатоцитами відокремлюють просвіт канальців від кровоносної системи печінки.
Канальцева мембрана містить транспортні білки для жовчних кислот, білірубіну, катіонів та аніонів. Мікроворсинки збільшують її площу. Органели представлені апаратом Гольджі і лізосомами. За допомогою везикул здійснюються транспорт білків (наприклад, IgA) від синусоїдальної до канальцевої мембрані, доставка синтезуються в клітці транспортних білків для холестерину, фосфоліпідів і, можливо, жовчних кислот з мікросом до канальцевої мембрані.
Цитоплазма гепатоцитів навколо канальці в додержіт структури цитоскелету: мікротрубочки, мікрофіламентиі проміжні філаменти.
Микротрубочки утворюються шляхом полімеризації тубуліну і формують мережу всередині клітини, особливо поблизу базолатеральной мембрани і апарату Гольджі, беручи участь в рецепторно-опосередкований везикулярного транспорті, секреції чи-пидов, а при певних умовах - і жовчних кислот. Освіта микротрубочек відзначено зниження колхицином.
У побудові микрофиламентов беруть участь взаємодіючі між собою полімерний (F) і вільний (G) актину. Мікрофіламенти, що концентруються навколо канальцевої мембрани, визначають скоротливу здатність і моторику канальців. Фаллоідін, що підсилює полімеризацію актину, і цітохалазін В, послабляє її, пригнічують моторику канальців і викликають холестаз.
Проміжні філаменти складаються з цитокератину і утворюють мережу між плазматичними мембранами, ядром, внутрішньоклітинними органелами і іншими структурами цитоскелету. Розрив проміжних філаментів призводить до порушення внутрішньоклітинних транспортних процесів і облітерації просвіту канальців.
Вода і електроліти впливають на склад канальцевого секрету, проникаючи через щільні контакти між гепатоцитами внаслідок осмотичного градієнта між просвітом канальців і просторами Діссе (Парацеллюлярний ток).Цілісність щільних контактів залежить від наявності на внутрішній поверхні плазматичної мембрани білка ZO-1 з молекулярної масою 225 кДа. Розрив щільних контактів супроводжується потраплянням в канальці розчинених більших молекул, що призводить до втрати осмотичного градієнта і розвитку холестазу. При цьому може спостерігатися регургітація канальцевої жовчі в синусоїди.
Жовчні канальці впадають в дуктул, звані іноді холангіол або каналами Герінга. Дуктул розташовуються в основному в портальних зонах і впадають в междольковие жовчні протоки, які першими з жовчних шляхів супроводжуються гілочками печінкової артерії і ворітної вени і виявляються в складі портальних тріад. Междольковие протоки, зливаючись, формують септальних протоки до тих пір, поки не утворюються два головних печінкових протока, що виходять з правої і лівої часток в області воріт печінки.
секреція жовчі
Освіта жовчі відбувається за участю ряду енергозалежних транспортних процесів. Її секреція відносно незалежна від перфузійного тиску. Сумарний струм жовчі у людини становить приблизно 600 мл / сут. Гепатоцити забезпечують секрецію двох фракцій жовчі: залежить від жовчних кислот ( «225 мл / добу) і не залежить від них (« 225 мл / добу). Решта 150 мл / сут секретируются клітинами жовчних проток.
Секреція солей жовчних кислот є найбільш важливим фактором освіти жовчі (Фракції, яка залежить від жовчних кислот).Вода рухається слідом за осмотически активними солями жовчних кислот. Зміна осмотической активності може регулювати надходження води в жовч. Є чітка кореляція між секрецією солей жовчних кислот і струмом жовчі.
Існування фракції жовчі, що не залежить від жовчних кислот, доведено можливістю утворення жовчі, яка не містить солей жовчних кислот. Таким чином, можливе продовження струму жовчі, незважаючи на відсутність екскреції солей жовчних кислот; секреція води при цьому обумовлена іншими осмотично активними розчинними речовинами, такими як глутатіон і бікарбонати.
Клітинні механізми секреції жовчі
Гепатоцит є полярну секреторну епітеліальну клітину, що має базолатеральную (синусоидальную і латеральну) і апикальную (канальцевую) мембрани.
Освіта жовчі включає в себе захоплення жовчних кислот та інших органічних і неорганічних іонів, транспорт їх через базолатеральную (синусоидальную) мембрану, цитоплазму і канальцеву мембрану. Цей процес супроводжується осмотической фільтрацією води, що міститься в гепатоците і парацеллюлярной просторі. Ідентифікація та характеристика транспортних білків синусоїдальної і канальцевої мембран складні. Особливо важким є вивчення секреторного апарату канальців, проте до теперішнього часу розроблена і довела свою надійність в багатьох дослідженнях методика отримання здвоєних гепатоцитів в короткоживущей культурі .. Клонування транспортних білків дозволяє охарактеризувати функцію кожного з них окремо.
Процес жёлчеобразованія залежить від "наявності певних білків-переносників в базолатеральной і канальцевої мембранах. Роль рушійної сили секреції виконує Na +, К + - АТФаза базолатеральной мембрани, забезпечуючи хімічний градієнт і різниця потенціалів між гепатоцитом і навколишнім простором. Na +, К + - АТФаза обмінює три внутрішньоклітинних іона натрію на два позаклітинних іона калію, підтримуючи градієнт концентрації натрію (висока зовні, низька всередині) і калію (низька зовні, висока всередині). В результаті цього вміст клітини має негативний заряд (-35 мВ) в порівнянні з позаклітинним простором, що полегшує захоплення позитивно заряджених іонів і екскрецію негативно заряджених іонів. Na +, К + -АТФаза не виявляється в канальцевої мембрані. Плинність мембран може впливати на активність ферменту.
Захоплення на поверхні синусоїдальної мембрани
Базолатеральную (синусоїдальна) мембрана має безліч транспортних систем для захоплення органічних аніонів, Субстратна специфічність яких частково збігається. Характеристика білків-переносників раніше давалася на підставі вивчення клітин тварин. Недавнє клонування транспортних білків людини дозволило краще охарактеризувати їх функцію. Транспортний білок для органічних аніонів (organic anion transporting protein - OATP) є натрійнезавісімим, переносить молекули ряду сполук, включаючи жовчні кислоти, бромсульфалеїн і, ймовірно, білірубін. Вважають, що транспорт білірубіну в гепатоцит здійснюють також інші переносники. Захоплення жовчних кислот, кон'югованих з таурином (або гліцином), здійснюється транспортним білком натрію / таурохолат (sodium / bile acid cotransporting protein - NTCP).
У перенесенні іонів через базолатеральную мембрану бере участь білок, обмінює Na + / H + і регулює рН всередині клітини. Цю функцію виконує також котранспортний білок для Na + / HCO 3 -. На поверхні базолатеральной мембрани відбувається також захоплення сульфатів, неетеріфіцірованних жирних кислот, органічних катіонів.
внутрішньоклітинний транспорт
Транспорт жовчних кислот в гепатоцитах здійснюється за допомогою цитозольних білків, серед яких основна роль належить За-гідроксістероіддегідрогеназе. Менше значення мають глутатіон-S-трансфераза і білки, що зв'язують жирні кислоти. У перенесенні жовчних кислот беруть участь ендоплазматичнийретикулум і апарат Гольджі. Везикулярний транспорт включається, мабуть, тільки при значному надходженні в клітину жовчних кислот (в концентраціях, що перевищують фізіологічні).
Транспорт білків рідкої фази і лігандів, таких як IgA і ліпопротеїди низької щільності, здійснюється за допомогою везикулярного трансцитозу. Час перенесення від базолатеральной до канальцевої мембрані становить близько 10 хв. Даний механізм відповідальний тільки за невелику частину сумарного струму жовчі і залежить від стану мікротрубочок.
канальцева секреція
Канальцева мембрана являє собою спеціалізовану дільницю плазматичноїмембрани гепатоцита, що містить транспортні білки (здебільшого АТФ-залежні), відповідальні за перенесення молекул в жовч проти градієнта концентрації. У канальцевої мембрані локалізуються також такі ферменти, як ЛФ, ГГТП. Перенесення глюкуронидов і глутатіон-S-кон'югатів (наприклад, білірубіну диглюкороніду) здійснюється за допомогою канальцевого мультіспеціфічного транспортного білка для органічних аніонів (сапаlicular multispecific organic anion transporter - cMOAT), перенесення жовчних кислот - за допомогою канальцевого транспортного білка для жовчних кислот (canalicular bile acid transporter - свати), функція якого частково управляється негативним внутрішньоклітинним потенціалом. Струм жовчі, що не залежить від жовчних кислот, визначається, по-видимому, транспортом глу-татіона, а також канальцевої секрецією бікарбонату, можливо, за участю білка, яка обмінює Cl - / HCO 3 -.
Важлива роль в транспорті речовин через канальцеву мембрану належить двом ферментам сімейства Р-глікопротеїнів; обидва ферменту є АТФ-залежними. Білок множинної медикаментозної резистентності 1 (multidrug resistance protein 1 - MDR1) переносить органічні катіони, а також здійснює виведення цитостатичних препаратів з ракових клітин, обумовлюючи їх резистентність до хіміотерапії (звідси назва білка). Ендогенний субстрат MDR1 невідомий. MDR3 переносить фосфоліпіди і діє як фліппаза для фосфатидилхоліну. Функція MDR3 і його важливе значення для секреції фосфоліпідів в жовч уточнені в експериментах на мишах, у яких відсутній mdr2-P-глікопротеїн (аналог MDR3 людини). При відсутності в жовчі фосфоліпідів жовчні кислоти викликають пошкодження біліарного епітелію, запалення дуктул і перідуктулярний фіброз.
Вода і неорганічні іони (особливо натрій) виводяться в жовчні капіляри по осмотичного градієнту шляхом дифузії через негативно заряджені напівпроникні щільні контакти.
Секреція жовчі регулюється багатьма гормонами і вторинними месенджера, включаючи цАМФ і протеїн С. Підвищення концентрації внутрішньоклітинного кальцію пригнічує секрецію жовчі. Пасаж жовчі по канальцям відбувається завдяки микрофиламентам, які забезпечують моторику і скорочення канальців.
дуктулярная секреція
Епітеліальні клітини дистальних проток виробляють збагачений бікарбонатами секрет, що модифікує складу канальцевої жовчі (так званий дуктулярная ток, жовчі).В процесі секреції відбувається вироблення цАМФ, деяких мембранних транспортних білків, включаючи білок, обмінює Cl - / HCO 3 -, і регулятор трансмембранної провідності при кістозному фіброзі -мембранний канал для Cl -, регульований цАМФ. Дуктулярная секреція стимулюється секретином.
Передбачається, що урсодезоксихолевая кислота активно всмоктується дуктулярная клітинами, обмінюється на бікарбонати, рециркулює в печінці і в подальшому знову виводиться у жовч ( «холегепатіческій шунт»). Можливо, цим пояснюється холеретичну дію урсодезоксихолевої кислоти, що супроводжується високою біліарної секрецією бікарбонатів при експериментальному цирозі.
Тиск в жовчних протоках, при якому відбувається секреція жовчі, в нормі становить 15-25 см вод. ст. Підвищення тиску до 35 см вод. ст. призводить до пригнічення секреції жовчі, розвитку жовтяниці. Секреція білірубіну і жовчних кислот може повністю припинятися, при цьому жовч стає безбарвною (Біла жовч)і нагадує слизову рідина.
Жовч є ізоосмотічная плазмі рідина, що складається з води, електролітів і органічних речовин (жовчних кислот, фосфоліпідів, холестерину, білірубіну). Жовчні кислоти (або їх солі) є основним органічним компонентом жовчі. Жовчні кислоти надходять в жовч з двох джерел: (1) первинні жовчні кислоти (холевая і хенодезоксихолева), які синтезуються з холестерину в печінці; (2) вторинні жовчні кислоти (дезоксихолева, літохолевая і урсодезоксихолевая) утворюються під дією бактерій кишечника з первинних жовчних кислот. Самі жовчні кислоти складаються з двох важливих компонентів, які і визначають їх фізіологічні та фізико-хімічні властивості: (1) стероїдного ядра з гідроксильними заступниками; (2) алифатической бічного ланцюга (рис. 7-4).
Мал. 7-4.
Жовчні кислоти складаються з двох компонентів - ядра з гідроксильними закінченнями і алифатической бічній ланцюжком. На малюнку холевая кислота показана як приклад трехгідроксільной кислоти (3? -, 7? -, 12? ОН). Іншими прикладами є жовчні кислоти, що містять дезоксихолат (З? -, 12? ОН), хенодезоксіхолатов (З? -, 7? ОН) і літохолат (3? ОН)
У більшості ссавців первинні жовчні кислоти містять від трьох до семи гідроксильних заступників, число яких впливає на їх водорастворимость (гідрофільність). Незабаром після утворення первинні жовчні кислоти піддаються модифікації в області термінальної карбоксильної групи. Це відбувається під час печінкової фази кишково-печінкової циркуляції вторинних жовчних кислот і кон'югацію їх з гліцином або таурином. Наявність гідрофільного (гідроксильні компоненти і амідні зв'язку алифатической бічного ланцюга) і гідрофобного (стероидное ядро) компонентів дозволяє кон'югованим молекулам жовчних кислот виступати в ролі амфотерного з'єднання. Це дає їм можливість формувати міцели (полімолекулярнимі агрегати) вище критичної міцелярної концентрації. У свою чергу, молекули жовчних кислот здатні розчиняти інші амфотерні речовини (холестерин, фосфоліпіди) з утворенням змішаних міцел. Ця детергентоподобная роль жовчних кислот важлива для стабілізації фізико-хімічного стану жовчі, перетравлення і всмоктування жирів.
Синтез жовчних кислот з холестерину регулюється за механізмом негативного зворотного зв'язку, хоча природа регуляції на молекулярному і біохімічному рівнях ще до кінця не з'ясована. Мікросомальне 7? -Гідроксілірованіе холестерину є критичним етапом в синтезі жовчних кислот. Хенодезоксіхоліевая кислота, яка використовується для розчинення каменів жовчного міхура, пригнічує синтез жовчних кислот і тим самим сприяє підвищенню рівня холестерину крові. При використанні ж урсодезоксихолевої кислоти таких змін не спостерігається навіть при тривалому лікуванні.
Освіта жовчі проходить як на синусоїдальної, так і на канальцевої поверхні мембрани гепатоцитів і є як інтрацелюлярна, так і парацеллюлярной процесом. На відміну від гломерулярної фільтрації в нирках, яка протікає пасивно під дією гідростатичних сил, в процесі утворення жовчі відбувається активне перенесення органічних і неорганічних компонентів в просвіт канальців і пасивний транспорт води. Таким чином, процеси секреції жовчі схожі з процесами секреції в ацинусах підшлункової залози, епітелії ниркових канальців. Освіта канальцевої жовчі можна розділити на два типи (рис. 7-5): (1) желчеобразование, залежне від секреції жовчних кислот, яке визначається як відношення кількості виділилася в канальці жовчі до кількості секретується солей жовчних кислот; (2) желчеобразование, незалежне від секреції жовчних кислот, яке може бути представлено як активна секреція неорганічних електролітів та інших речовин і відображено на графіку у вигляді y-перетину цієї лінії. Іншими словами, пов'язане з секрецією кислот желчеобразование є швидкість потоку жовчі, що залежить від наявності в жовчних канальцях осмотично активних жовчних солей, а несвязанное з секрецією кислот желчеобразование - при відсутності солей жовчних кислот. Ставлення швидкості утворення жовчі і утворення солей жовчних кислот має нелінійний характер при невеликих кількостях виділяється жовчі і не може відповідати лінійної залежності, показаної на Рис. 7-5. Тому обидва види желчеобразования повинні розглядатися як взаємозалежні показники утворення жовчі.
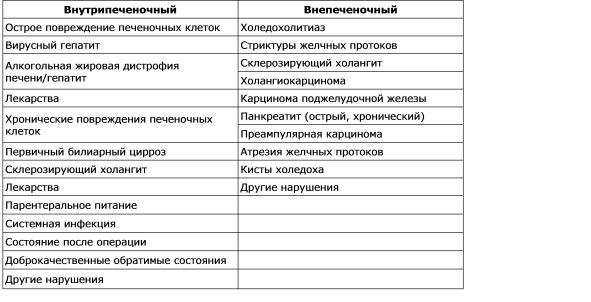
Порушення утворення жовчі називається холестазом. Послідовність виникають патологічних, фізіологічних і клінічних проявів холестазу залежить від його причини. При морфологічному дослідженні матеріалу гепатобіопсій жовч виявляється в канальцях періцентральная гепатоцитів, відзначається дилатація канальців, а при дослідженні ультраструктури виявляється зниження кількості мікроворсинок. Холестаз можна визначити як функціональний дефект утворення жовчі на рівні гепатоцита (внутрішньопечінковий холестаз), а також як органічні або фізичні проблеми секреції і відтоку жовчі (позапечінкові холестаз). Найбільш часті причини внутрішньо-і внепеченочного холестазу наведені в табл. 7-2. Існує кілька механізмів, що грають важливу роль в патогенезі внутрішньопечінкового холестазу: порушення і пошкодження функції синусоїдальної мембрани; порушення функцій внутрішньоклітинних органел гепатоцитів; ушкодження і порушення функцій канальцевої мембрани. Таким чином, не існує єдиного механізму холестазу в різних клінічних ситуаціях, а численність механізмів може призводити до різних розладів. Клінічно холестаз характеризується підвищенням в крові рівня багатьох речовин, включаючи білірубін, солі жовчних кислот, холестерин, які в нормі секретуються в жовч. При біохімічному аналізі крові при холестазі спостерігається непропорційний підвищення активності лужної фосфатази і концентрації білірубіну паралельно зі змінами рівня амінотрансфераз, про які йтиметься нижче.
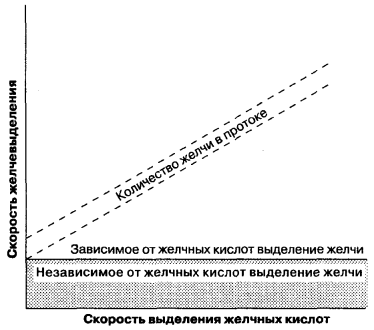
Мал. 7-5.
Жовчоутворення, пов'язане з секрецією жовчних кислот, желчеобразование, незалежне від секреції жовчних кислот. (За: Moseley R. H., Bile secretion. In: Yamada Т., Alpers D. H., Owyang C., Powell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995: 387.)
Таблиця 7 - 2.
У печінці утворюється найважливіший травний сік - жовч.
Жовч виробляється гепатоцитами шляхом активного і пасивного транспорту в них води, холестерину, білірубіну, катіонів. В гепатоцитах з холестерину утворюються первинні жовчні кислоти - холевая і дезоксихолева. З білірубіну і глюкуронової кислоти синтезується водорозчинний комплекс. Вони надходять в жовчні капіляри і протоки, де жовчні кислоти з'єднуються з гліцином і таурином. В результаті утворюються глікохолевой і таурохолевая кислоти. Гідрокарбонат натрію утворюється за допомогою тих же механізмів, що і в підшлунковій залозі.
Жовч виробляється печінкою постійно. У добу її утворюється близько 1 літра. Гепатоцитами виділяється первинна або печінкова жовч. Це рідина золотисто-жовтого кольору лужної реакції. Її рН = 7,4 - 8,6. Вона складається з 97,5% води і 2,5% сухого залишку. У сухому залишку містяться:
1. Мінеральні речовини. Катіони натрію, калію, кальцію, гідрокарбонат, фосфат аніони, аніони хлору.
2.Желчние кислоти - таурохолевая і глікохолевой.
3.Желчние пігменти - білірубін і його окислена форма биливердин. Білірубін надає жовчі колір.
4.Холестерін і жирні кислоти.
5.Мочевіна, сечова кислота, креатинін.
Оскільки поза травлення сфінктер Одді, розташований в гирлі загальної жовчної протоки, закритий, що виділяється жовч накопичується в жовчному міхурі. Тут з неї реабсорбується вода, а зміст основних органічних компонентів і муцина зростає в 5-10 разів. Тому міхурово жовч містить 92% води і 8% сухого залишку. Вона більш темна, густа і в'язка, ніж печінкова. Завдяки цій концентрації міхур може накопичувати жовч протягом 12 годин. Під час травлення відкривається сфінктер Одді і сфінктер Люткенса в шийці міхура. Жовч виходить в дванадцятипалу кишку.
Значення жовчі:
1.Желчние кислоти емульгують частина жирів, перетворюючи великі жирові частинки в дрібнодисперсні краплі.
2.Она активує ферменти кишкового і панкреатичного соку, особливо ліпази.
3.В комплексі з жовчними кислотами відбувається всмоктування довголанцюгових жирних кислот і жиророзчинних вітамінів через мембрану ентероцитів.
4.Желчь сприяє ресинтезу тригліцеридів в ентероцитах.
5.Інактівірует пепсину, а також нейтралізує кислий химус, що надходить зі шлунка. Цим забезпечується перехід від шлункового до кишковому травленню.
6.Стімулірует секрецію підшлункового і кишкового соків, а також проліферацію і злущування ентероцитів.
7.Усілівает моторику кишечника.
8.Оказивает бактеріостатичну дію на мікроорганізми кишечника і таким чином перешкоджає розвитку гнильних процесів в ньому.
Регуляція жовчоутворення і жовчовиділення в основному здійснюється гуморальними механізмами, хоча деяку роль відіграють і нервові. Найпотужнішим стимулятором желчеобразования в печінці є жовчні кислоти, які всмоктуються в кров з кишечника. Його також підсилює секретин, який сприяє збільшенню вмісту в жовчі бікарбонату натрію. Блукаючий нерв стимулює вироблення жовчі, симпатичні гальмують.
При надходженні хімусу в дванадцятипалу кишку починається виділення I-клітинами її слизової холецистокініну-панкреозимина. Особливо цей процес стимулюють жири, яєчний жовток і сульфат магнію. ХЦК-ПЗ посилює скорочення гладких м'язів міхура, жовчних проток, але розслаблює сфінктери Люткенса і Одді. Жовч викидається в кишку. Рефлекторні механізми відіграють невелику роль. Химус дратує хеморецептори тонкого кишечника. Імпульси від них надходять в травний центр довгастого мозку. Від нього вони по вагусу до жовчовивідних шляхах. Сфінктери розслабляються, а гладкі м'язи міхура скорочується. Це сприяє желчевиведеніе.
Найбільш важкими захворюваннями є гепатити та цироз печінки. Найчастіше гепатити є наслідком інфекції (інфекційні гепатити А, В, С) і впливу токсичних продуктів (алкоголь). При гепатитах уражаються гепатоцити і порушуються всі функції печінки. Цироз це результат гепатитів. Найчастішим порушенням жовчовиділення є жовчнокам'яна хвороба. Основна маса жовчних каменів утворена холестерином, так як жовч таких хворих перенасичена Ними.
Печінка - це найбільший орган тіла, що займає центральне місце в обміні речовин. Вона виконує безліч функцій, беручи участь в обміні білків, вуглеводів, жирів, гормонів і вітамінів, а також в знешкодженні багатьох ендогенних і екзогенних речовин. Ці процеси описані в підручниках фізіологічної хімії, а в цьому розділі в зв'язку з травною системою ми розглянемо лише видільну функцію печінки,тобто секрецію жовчі. жовч складається з води, мінеральних солей, слизу, ліпідів холестеролу і лецитину і двох видів специфічних компонентів - жовчних кислот і пігменту білірубіну. Жовчні кислоти є детергентами, і їх емульгуючу дію грає важливу роль у перетравленні ліпідів. Білірубін - це кінцевий продукт розпаду гемоглобіну, що підлягає виведенню з організму.
^
Освіта жовчі
Функціональна анатомія.клітини печінки (Гепатоцити)утворюють пластинки товщиною в одну клітку, розділені вузькими щілинами (Простір Діссе),що представляють собою заповнені
^ 766 ЧАСТИНА VIII. ХАРЧУВАННЯ, ТРАВЛЕННЯ І ВИДІЛЕННЯ
кров'ю синусоїди,еквівалентні капілярах. У стінках синусоїдів є пори, через які можуть проходити такі великі макромолекули, як альбумін і ліпопротеїни. за дрібним канальцам-жовчним капілярах, обмеженим плазматическими мембранами двох сусідніх гепатоцитів, - жовч збирається в більші канальці Герінга,стінки яких, як і більших за розміром междол'кових канальціві жовчних проток,утворені кубічними секреторними клітинами. Дрібні канальці всередині часточок печінки і між ними зливаються в більші, утворюючи в кінцевому рахунку печінковий протік.Від цього протока відходить міхуровопротокадо жовчного міхура. Після злиття печінковий і міхурово протоки утворюють загальний жовчний протік,відкривається в дванадцятипалу кишку на вершині фатерова сосочка позаду протоки підшлункової залози або поруч з ним (рис. 29.1).
^ Функції жовчі.Жовч виконує безліч важливих функцій. Разом з нею виводяться кінцеві продуктиобміну, наприклад білірубін, а також лікарські препарати і токсини. виділення зжовчю холестеролувідіграє важливу роль в регуляції його балансу. жовчні кислотинеобхідні для емульгування і всмоктування жирів. Крім того, жовч містить воду, мінеральні солі та слиз. У добу виділяється близько 600 млжовчі, і 2/3 цієї кількості надходить з канальців, а 1/3-з більших проток.
^ канальцева жовч утворюється приблизно в рівних кількостях за участю двох різних механізмів-залежного від жовчних кислот і незалежного від них (рис. 29.27).
^ Секреція, залежна від жовчних кислот. Встановлено тісний взаємозв'язок між швидкістю виділення жовчіі секрецією жовчних кислот.У канальцевої жовчі концентрація жовчних кислот в 100 разів вище, ніж в портальної крові, тому вважають, що вони виділяються шляхом активного транспорту за участю переносника.Слідом за жовчними кислотами по осмотичного градієнту в канальці спрямовується вода, тому жовч Ізотонічність крові.
Існує два джерела жовчних кислот. По-перше, вони синтезуються в самих гепатоцитах de novo з холестеролу за участю 7-гідроксилази. Даний фермент відіграє ключову роль і за механізмом зворотного зв'язку контролює швидкість синтезу жовчних кислот. По-друге, гепатоцити здатні активно поглинати жовчні кислоти з портальної кровіі виділяти їх в канальці (див. також рис. 29.29). Ця екстракція дуже ефективна; при одноразовому проходженні крові через печінку з неї витягується 80% жовчних кислот. Тому
Концентрація жовчних кислот в периферичної крові набагато нижче, ніж в ворітної системі. Оскільки жовчні кислоти витягуються з крові в 6 разів швидше, ніж надходять в канальці, саме останній процес лімітує швидкість секреції жовчних кислот.
^ Секреція, незалежна від жовчних кислот. В
Цьому процесі беруть участь іони Na +, Cl -, HCO 3 - і вода. Рушійною силою служить активний транспорт Να + , Можливо разом з бікарбонатом. Секрецію, незалежну від жовчних кислот, стимулює, зокрема, секретин.
Крім жовчних кислот в канальці активно секретуються білірубін, холестерол і фосфоліпіди (переважно лецитин) (рис. 29.27). Нерозчинний у воді ( «Непрямий») білірубін,велика частина якого утворюється з гемоглобіну постарілих еритроцитів, надходить в гепатоцити у вигляді колоїдного агрегату, пов'язаного з альбуміном. Добове освіту його становить близько 4 г / кг маси тіла, або 200-300 мг / сут. В гепатоцитах 80% білірубіну Кон'юговані зглюкуроновоюкислотою і невелика його кількість - з сірчаною кислотою. У такій кон'югованої
^ ГЛАВА 29. Функції шлунково-кишкового ТРАКТУ 767
Формі білірубін виділяється з жовчю ( «Прямий» білірубін).В основному тим же шляхом виводяться лікарські препарати і токсини.
^ Модифікація жовчі в жовчних протоках
(Рис. 29.27). У протоках, куди відкриваються канальці, первинна жовч піддається модифікації. Цей процес нагадує процес модифікації клубочкового фільтрату в ниркових канальцях (с. 785), і подібним чином для нього розраховується кліренс,тільки в разі жовчі в якості інертного речовини використовують замість інуліну ерітрітолабо манітол,які секретуються в канальці, але не реабсорбуються. Подібні дослідження показали, що близько 180 мл жовчі, або 1/3 її загальної кількості, виділяється в протоки при активній секреції НСО ^ ". Цей процес стимулюється секретином.
^
Печінкова і міхурово жовч
Склад печінкової жовчі(Табл. 29.3). Жовч, що виділяється печінкою зі швидкістю 0,4 мл / хв, має золотистий колір, що пояснюється присутністю в ній білірубіну. Концентрація електролітів в цій жовчі така ж, як в плазмі, за винятком того, що вона містить в два рази більше НСОз "і дещо менше С1 ~. У той же час за складом органічних речовин жовч сильно відрізняється від плазми, так як в жовчі вони представлені майже виключно жовчними кислотами, холестеролом і фосфоліпідами.
 |
^ Жовчні кислотиутворюються в печінці з холестеролу в результаті його гідроксилювання і приєднання карбоксильної групи. Утворені в печінці кислоти - це первинні жовчні кислоти;до них відносяться хенодезоксихолева(Діоксіформа) і холевая(Тріоксіформа) кислоти.У печінці вони знаходяться не у вільній формі, а у вигляді кон'югатів з гліцином і таурином, причому кон'югатів з гліцином утворюється в три рази більше, оскільки кількість таурину обмежена. Кон'юговані жовчні кислоти краще розчинні у воді,
Чим некон'югірованная, і мають більшу здатність до дисоціації і утворенню жовчних солей з катіонами, головним чином з іонами Na +. У кислому середовищі (pH 4,0) солі жовчних кислот нерозчинні і випадають в осад, але при фізіологічних значеннях pH (в тонкому кишечнику) вони добре розчинні.
У дисталь відділі клубової кишки і в товстій кишці частина солей первинних жовчних кислот піддається дегідроксілірованію під дією анаеробних бактерій і перетворюється у вторинні жовчні кислоти-литохолевую(Монооксіформа) і дезоксихолевої(Діоксіформа). Хенодезоксихолева, холевая і дезоксихолева кислоти присутні в співвідношенні 2: 2: 1. Литохолевая кислота присутня лише в деяких фракціях, оскільки велика її частина екскретується.
Емульгуючу дію жовчних кислот на жири засновано головним чином на їх здатності утворювати міцели.Молекули жовчних кислот мають таку тривимірну структуру, при якій гідрофільні карбоксильні і гідроксильні групи знаходяться на одній стороні молекули, а гідрофобна частина молекули (стероидное ядро, метильние групи) -на протилежної, за рахунок чого молекули жовчних кислот володіють і гідрофільними,і ліпофіл'нимі властивостями.Завдяки такій будові молекули жовчних кислот діють як детергенти: на кордоні розділу ліпідної і водної фаз вони утворюють майже мономолекулярну плівку, в якій гідрофільні групи звернені до водної, а ліпофільні-к ліпідної фазі. У водному середовищі жовчні кислоти утворюють впорядковані агрегати -міцеллиза умови, що їх концентрація досягає певного рівня, званого критичної концентрацією міцелоутворення(1-2 ммоль / л). Внутрішня, липофильная область міцели може містити ліпіди,наприклад холестерол і фосфоліпіди; такі міцели називають змішаними (рис. 29.28). Сам по собі холестерол не розчиняється у воді, але в складі міцел може перебувати в розчині. Якщо його концентрація перевищує ємність мицелл, він утворює кристалічний осад; цей процес лежить в основі освіти холестеролових жовчних каменів (с. 769).
^ Склад міхурово жовчі (Табл. 29.3). Ємність жовчного міхура складає всього 50-60 мл. У той же час печінку секретирует жовч зі швидкістю 600 мл / сут, і половина цієї кількості перед надходженням в тонкий кишечник проходить через жовчний міхур. Різниця між обсягом жовчі, що надходить в жовчний міхур, і його ємністю компенсується високоефективної реабсорбциейводи в жовчному міхурі. Протягом декількох годин з жовчі може реабсорбироваться 90% води. при
^
768 ЧАСТИНА VIII. ХАРЧУВАННЯ, ТРАВЛЕННЯ І ВИДІЛЕННЯ
це органічні речовини залишаються в жовчному міхурі і їх концентрація в жовчі підвищується. Рушійною силою реабсорбції служить активний транспорт іонів Nα *за участю «насоса», вбудованого в базальну і латеральну мембрани клітин і активується (Na + -K +) - ATФазой. Слідом за іонами Na + переміщаються іони Cl - і HCO 3 -, диффундирующие в напрямку електричного градієнта або транспортуються переносниками. В результаті реабсорбції HCO 3 - pH міхурово жовчі зменшується до 6,5 проти 8,2 в печінкової жовчі. Внаслідок створення в міжклітинному просторі епітелію жовчного міхура високої концентрації іонів Na + виникає осмотичний градієнт, що приводить до накачиванию води, яка відтікає потім в капіляри (с. 751).
^ Моторика жовчного міхура. У стані натщесерце жовч накопичується в жовчному міхурі, а у
Час прийому їжі виділяється в результаті скорочень жовчного міхура. Основним стимулятором скоротливої активності жовчного міхура служить холецистокінін,секретується слизовою оболонкою дванадцятипалої кишки при вступі до неї хімусу, що містить жири. До деякої міри скорочення жовчного міхура стимулюються також блукаючим нервомі парасімпатолітікамі.Вони починаються вже через 2 хв після того, як жирна їжа стикається зі слизової кишечника, і через 15-90 хв міхур повністю спорожняється. Моторика жовчного міхура включає два процеси. Спочатку розвивається тонічне скорочення, внаслідок чого зменшується діаметр жовчного міхура, а потім на цей ефект накладаються періодичні скорочення, частота яких становить 2-6 / хв. В результаті цих двох процесів створюється тиск 25-30 мм рт. ст.
^
Кишково-печінкова циркуляція
Циркуляція жовчних кислот(Рис. 29.29). Жовчні кислоти виділяються в дванадцятипалу кишку у вигляді змішаних міцел. Незважаючи на розведення жовчних кислот вмістом шлунку, їх концентрація в кишечнику становить близько 10 ммоль / л і залишається вище критичної концентрації міцелоутворення. Тут на додаток до холестеролу і лецитину в міцели включаються продукти гідролітичного розщеплення жирів - жирні кислотиі моногліцериди.При первинному контакті мицелл з кишковою стінкою
^
ГЛАВА 29. Функції шлунково-кишкового ТРАКТУ 769
Ліпіди дифундують через мембрану щіткової облямівки в ентероцита, а жовчні кислоти залишаються в просвіті кишечника, але при подальшому проходженні по кишечнику жовчні кислоти всмоктуються шляхом активного і пасивного транспорту.
Близько 50% жовчних кислот всмоктується в кишечнику пасивним шляхом. В результаті розщеплення кон'югатів жовчних кислот і дегідроксілірованія останніх під дією кишкових бактерій підвищується їх розчинність в ліпідах і полегшується пасивна дифузія.
^ активне всмоктування жовчних кислот відбувається виключно в кінцевому відділі клубової кишки-рідкісне явище, відоме ще тільки для всмоктування вітаміну В12. Активному всмоктуванню піддаються лише ті жовчні кислоти, які мають великий полярністю, що утрудняє їх пасивне всмоктування, наприклад кон'югати таурину. Для процесу всмоктування жовчних кислот у кінцевому відділі клубової кишки характерні типові ознаки активного транспорту: кінетика насичення і конкурентне інгібування. Невелика кількість жовчних кислот (7-20%) не включається ні в активну, ні в пасивне всмоктування і виводиться з організму.
Присутність жовчних кислот в товстій кишці відіграє, мабуть, важливу роль в регуляції консистенції калових мас. При концентрації діоксіжелчних кислот в товстій кишці понад 3 ммоль / л в просвіт кишечника секретується значна кількість електролітів і води, що призводить до проносу. Різко виражена форма цього «Хологенная» проносуможе спостерігатися при резекції або захворюванні кінцевого відділу клубової кишки, і для його лікування використовують зв'язування жовчних кислот за допомогою іонообмінника холестираміну.
При надходженні поглинених жовчних кислот в печінкуз них знову утворюються кон'югати, а деякі вторинні жовчні кислоти піддаються гидроксилированию. Втрата жовчних кислот з калом (02-06 г / день) компенсується за рахунок їх синтезу.
^ Загальний пул жовчних кислот в організмі становить близько 3,0 м Цієї кількості недостатньо для забезпечення ліполізу після прийому їжі; зокрема, при споживанні жирної їжі жовчних кислот потрібно в 5 разів більше. Однак організм не відчуває дефіциту в жовчних кислотах, оскільки вони багаторазово циркулюють через кишечник і печінку (Кишково-печінкова циркуляція).Частота, з якою пул жовчних кислот робить повний цикл, залежить від харчового режиму і становить від 4 до 12 циклів на добу.
^ Циркуляція білірубіну. жовчний пігмент білірубін,подібно жовчним кислотам і ліпідів, надходить в кишечник в формі глкжуроніда. Лише невелика кількість цього полярного з'єднання
Реабсорбується в жовчному міхурі і тонкому кишечнику. У кінцевому відділі клубової кишки і (в основному) в товстій кишці кон'югат білірубіну розщеплюється під дією бактеріальних гидролаз. При цьому білірубін перетворюється в уробилиноген,який поряд з іншими продуктами розпаду білірубіну надає коричневий колір калових мас. Менше 20% уробилиногена всмоктується назад, і з цієї кількості близько 90% знову потрапляє в печінку і повертається в жовч, а решта 10% виводяться з сечею.
Патофізіологічні аспекти. Підвищений вміст уробіліногену в сечі може вказувати на захворювання печінки,що супроводжується порушенням екскреції білірубіну. Повна відсутність уробіліногену в сечі, світлий колір калу і жовтушність вказують на повну закупорку жовчного міхура;в цій ситуації білірубін зовсім не надходить в кишечник і уробіліноген не утворюється.
Найбільш відоме і поширене порушення нормальної фізіології жовчної системи-це випадання в осад холестсрола з утворенням холестеролоаих жовчних каменів. Холестерол, як і лецитин, знаходиться в розчиненому стані тільки в складі змішаних міцел. Якщо зростає концентрація xo.jecmepo.ia або стає нижче критичного рівні концентрація жовчних кислот або лецитину,холестерол випадає в осад. До числа факторів, що викликають підвищення відносного вмісту холестеролу, відносяться естрогени, вуглеводна дієта, надлишкова маса і процеси, що знижують концентрацію жовчних кислот, наприклад запалення клубової кишки (Хвороба Крона)або її резекція. У деяких випадках досить буває перорального прийому жовчних кислот, щоб літогенні жовч перетворилася в алітогенную, в якій холестеролових камені можуть розчинитися. Найбільше для цієї мети підходять хенодезоксихолева і уродезоксіхолевой кислоти, так як вони не викликають проносу.
Клінічним проявом порушеного обміну білірубіну є жовтяниця. Жовтий колір шкіри пов'язаний з підвищеним вмістом в плазмі білірубіну, яке може мати місце в наступних випадках:
при підвищеному утворенні білірубіну в результаті посиленого розпаду еритроцитів (гемолітична жовтяниця);
в результаті порушення процесу кон'югированія або транспорту білірубіну в гепатоцитах, як, наприклад, при жовтяниці вагітнихабо пологової жовтяниці Джильберта;
при затримці відтоку жовчі, наприклад через жовчних каменів або пухлин, локалізованих в області жовчної протоки (механічна жовтяниця).