Epe kialakulása. Az epe képződésének fiziológiája. Gall formáció
Az epe kialakulása a májban a vérben, a sejtekben és a sejtsejteken keresztül keringő anyagok (víz, glükóz, kreatinin, elektrolitok, hormonok, vitaminok stb.) Aktív és passzív szállításával, valamint az epe komponensek (epesavak) hepatociták által történő aktív szekréciójával következik be. és a víz és egyes anyagok kisbélcsatornákból és epekövekből való újbóli felszívódása. Ennek a folyamatnak a fiziológiai jelentősége változatos. Az epe elsődlegesen emésztőrendszernek tekinthető, mivel az epesavak (főként szerves anionjaik) kulcsszerepet játszanak a zsírok felszívódásában. Az epe emulgeálja a zsírokat, növelve a felületet, amelyen a lipáz hatására hidrolizálódnak. Az epe hatására a zsírhidrolízis termékek oldódnak. Ez elősegíti a trigliceridek felszívódását és reszintézisét enterocitákban. Az epe növeli a hasnyálmirigy és a bél enzimek (különösen lipázok) aktivitását, fokozza a fehérjék és szénhidrátok hidrolízisét és felszívódását.
A zsírok emésztése megsérti a rosszul emésztett és más élelmiszereket, mivel a zsír az élelmiszer legkisebb részeit fedi le, és megakadályozza az enzimek hatását. Ilyen körülmények között a bélbaktériumok aktivitása fokozott bomlási folyamatokat, fermentációt és gázképződést eredményez.
Szabályozási hatások is jellemzőek az epe - az epe képződésének stimulálása, az epe kiválasztása, a vékonybél motoros és szekréciós aktivitása, valamint az enterociták proliferációja és desquamációja. Az epe gátolja a gyomor emésztésének folyamatát a duodenumba belépő sav semlegesítésével és a pepszin inaktiválásával, előkészítve a bélben történő emésztésre. Szintén szabályozási hatással van a gastroduodenalis komplex evakuációs tevékenységére. Fontos az epe szerepe a zsírban oldódó vitaminok (A, D, E és K) felszívódásában, a koleszterin, a béllumenből származó kalcium sókban.
Ezen túlmenően az epe képződése és szekréciója bizonyos molekulák és ionok különös kiválasztási módjának tekinthető, amelyet a vesén keresztül nem lehet kiválasztani. Ezek közül a legjelentősebb a koleszterin (a szabad koleszterin, az észterek és az epesavak), a bilirubin, valamint a réz és vas molekulák. Ezért az epe kiválasztási folyadéknak tekintendő.
Az epe 80% vizet és 20% -ban feloldott anyagot tartalmaz. Az utóbbiak közé tartoznak az epesavak és ezek sói (kb. 65%), a biliáris foszfolipidek (körülbelül 20%, főként lecitin miatt), fehérjék (kb. 5%), koleszterin (4%), konjugált bilirubin (0,3%), enzimek. , immunglobulinok, valamint számos, epe által kiváltott exogén és endogén anyag (növényi sztirolok, vitaminok, hormonok, gyógyszerek, toxinok, fémionok - réz, vas, kálium, nátrium, kalcium, cink, magnézium, higany stb.). Átlagosan 600-700 ml epe szekretálódik naponta (250 - 1500 ml, kb. 10,5 ml / 1 kg testtömeg). Ugyanakkor körülbelül 500 ml / nap a teljes térfogatot a hepatociták szekréciója és körülbelül 150 ml / nap adagolja az epevezetékek sejtjeinek szekretálása.
Epe kialakulása (Cholepoiesis) folyamatosan megy, és az epe áramlása a nyombélbe (Holekinez) rendszeresen történik. Üres gyomorban az epe a bélbe kerül az időszakos éhes aktivitásnak megfelelően. A pihenőidők alatt a WB-be megy, ahol koncentrálódik, valamivel megváltoztatja összetételét és letétbe helyezi. A víz és a sók mellett a koleszterin és a szabad zsírsavak is felszívódnak. Ebben a tekintetben különbséget kell tenni a máj és az epehólyag epével.
Az epe kis enzimatikus aktivitással rendelkezik; A hepatikus epe pH-értéke 7,3-8,0. A béltartalommal ellentétben szinte semmilyen baktériumot nem tartalmaz. Az epe sterilitását biztosító tényezők közé tartozik az epesavak jelenléte (bakteriosztatikus hatás), az immunglobulinok gazdag tartalma, a nyálelválasztás, az epeenergia-szubsztrátok relatív szegénysége a baktériumok számára.
Az Epe egy micelláris oldat. A koleszterin, amely gyakorlatilag nem oldódik vízben, oldott állapotban szállítható az epében, a micelláris szerkezete miatt. Ezt az eljárást kolloid oldódásnak - szolubilizációnak nevezik.
Az epesavak olyan felületaktív, amfipatikus (mind hidrofób és hidrofil) molekulák, amelyek képesek ön-aggregálódásra. Ebben az esetben egy nagyon szűk koncentrációs sebességnek, a miklizáció kritikus koncentrációjának köszönhetően egyszerű micellák képződnek. Az egyszerű micellák kifejezetten képesek a lipidek feloldására, vegyes micellákat képezve.
A vegyes micelláknak feltételezhetően hengeres szerkezetűek: a hengeres szár poláris lipidekkel van feltöltve, és az epesav molekulák a lipidmolekulák poláris végei között vannak, amelyek hidrofil oldallal néznek a vízkörnyezet felé, ami meghatározza a hidrofilitást (vízoldhatóságot).
A vegyes micellák olyan esszenciális komponenseket tartalmaznak, amelyek kívül eső epesavak, foszfolipidek (főként foszfatidil-kolin - lecitin) és koleszterin találhatóak a micellában.
Az epe színe sárgásbarna a bilirubin jelenléte miatt, amelynek legnagyobb része a diglukuronid bilirubin, annál kisebb - monoglukuronid bilirubin formájában - formájában jelenik meg. A társult bilirubin nem része a micelláknak.
Mivel az intercelluláris epehólyag fa vízáteresztő, az epehólyag és a máj epe izotóniás.
Epe kialakulása
Biliáris szekréció
Az epesavak a biliáris szekréció fő összetevői, csak a májban képződnek. Elsődleges epesavak - koleszterin hepatocitákban szintetizálódik a trihidroxi-cholisz (cholium) és a dihidroxi-chols (chenodeoxycholic). Másodlagos epesavak (dezoxikolsav és kis mennyiségben - litokolikus) a vastagbélben keletkezik primerből a nukleáris hidroxilcsoportok bakteriális módosítása következtében (7a-dehidroxilezés anaerob baktériumok szabályozása alatt). Tercier epesavak (főként ursodeoxikol) képződik a májban szekunder epesavak izomerizálásával.
A koleszterinből származó epesavak szintézisét a negatív visszacsatolás mechanizmusa szabályozza: az epesavaknak a portálvérrel rendelkező hepatocitákhoz való csökkentett visszatérése bioszintézisük növekedéséhez vezet; az epesavak szintézisének bármilyen növekedése a koleszterin képződésének megfelelő növekedésével jár.
Az epesavak a májban a glicin aminosavakkal (kb. 80%) vagy taurinnal (kb. 20%) konjugálódnak. A szénhidrátban gazdag étel fogyasztásakor a glikokolsav mennyisége nő az epében, és a magas fehérjetartalmú étrendben a glikofenolsavak mennyisége nő. Az aminosavkötés megakadályozza az epesavak felszívódását az epehólyagban és a vékonybél kezdeti szakaszaiban (csak a terminális ileumban és a vastagbélben). Baktériumok hatására az epesók hidrolízise az epesavak, glicin vagy taurin kialakulásával lehetséges.
Az epesavak bioszintézise után, amint azt fentebb említettük, karboxilcsoportjuk a glicin vagy a taurin aminocsoportjához kapcsolódik. Kémiai szempontból egy ilyen kötés egy gyenge savat erősebbé alakít. A megkötött epesavak alacsony pH-értéken leginkább oldódnak és a kalciumionokkal (Ca 2+) szemben a kicsapódás ellenállóbbak, mint a nem kötött epesavak. Biológiai szempontból a kötési eljárás nem teszi lehetővé az epesavakat a sejtmembránokon keresztül történő penetrációra. Ezért az epevezetékekben vagy a vékonybélben abszorbeálódnak, hordozómolekulák jelenlétében, vagy ha az epesavak bakteriális hasításon mennek keresztül. E szabály alól kivétel a glicin-dihidroxi-kapcsolódó epesavak, mivel passzívan abszorbeálhatók, ha hidrogénionot (H +) kapnak.
Az epesavak nagy része (kb. 85-90%), melyet az epe áramlása a vékonybélbe áramol, bejut a vérbe. Ugyanakkor a vékonybélbe szekretált legtöbb epe-sav felszívódik sértetlenül. A sav kisebb része nem szívódik fel, mivel bakteriális hasításon megy keresztül a disztális vékonybélben. Ezek passzívan felszívódnak, és a májba visszatérve ismét kötődnek és szekretálódnak az epébe. Az epesavak fennmaradó 10-15% -a elsősorban a székletből kerül ki a szervezetből. Ezt az epesav-veszteséget kompenzálja a hepatociták szintézise.
Ez a bélbontási folyamat és a májban való reabszorpció normális része az epesavak metabolizmusának. A nem felszívódó epesavak kis része belép a vastagbélbe. Itt befejeződik a hasítási folyamat. Ezenkívül másodlagos epesavak képződnek a vastagbélben anaerob baktériumok hatására (lásd fent).
A dezoxikolsav és a lithocholic savak részlegesen felszívódnak a vastagbélben és visszatérnek a májba. A visszatérés után az epesavak metabolizmusa más. A dezoxikolsav glicinhez vagy taurinhoz kötődik, és a primer epesavakkal kering. Meg kell jegyezni, hogy az időseknél az epe összetételében a dezoxikolsav a fő epesav. A litokolsav nem csak a glicinhez vagy a taurinhoz kötődik, hanem a C-3 pozícióban szulfátokhoz is. Az ilyen "kettős" kötés csökkenti a bélben való felszívódás lehetőségét, aminek következtében a keringő epesavak összetételéből a lithocholic sav gyorsan elveszik - az epe tartalma ritkán haladja meg az 5% -ot. A legtöbb (95%) epesav, amely a felnőttek epe része, a cholus, chenodesoxycholic és deoxycholic acid.
Az epesavaknak a hepatocitákban való kötésének folyamata nagyon hatékony, így a biliáris epesavak teljesen konjugált formában vannak jelen. Az ezt követő bakteriális hasítás és a dehidroxilezés azt a tényt eredményezi, hogy a székletben az epesavak nem kötődnek.
A fent leírt primer epesavak dehidroxilezésének folyamata csökkenti a vízben való oldódási képességüket. Másodlagos epesavak képződése emberben gyakorlatilag nincs fiziológiai jelentősége, ellentétben az állatokkal. A deoxikolsav túlzott felszívódása a vastagbélben növeli a koleszterin-epekő kockázatát. A litokolsavat hepatotoxikusnak tekintik. Az állatkísérletek kimutatták, hogy a lithocholic acid felhalmozódása a májban a vereséghez vezet. Azonban még nem bizonyították, hogy ennek a savnak az emberben történő felszívódásának növekedése a máj megzavarásához vezet.
Az ursodeoxikolsav, valamint a deoxikolsav a májban kötődik, és az elsődleges epesavakkal kering. Ennek a savnak a metabolikus útja azonban nagyon rövid, és a kötésben lévő ursodeoxikolsav tartalma az epe soha nem haladja meg az epesavak teljes mennyiségének 5% -át. Úgy véljük, hogy az ursodeoxikolsav képződése nem fontos fiziológiai jelentőséggel bír.
Az epesavak a zsírok erős oldószerei, így azok citotoxikusak olyan koncentrációkban, amelyek elérik a miklizáció kritikus koncentrációját. Ugyanakkor az epeben lévő kötődő chenodeoxikol és deoxikolsavak sokkal toxikusabbak, mint a cholic és ursodeoxycholic. Bár az epesavak in vitro demonstratív módon citotoxikusak, az epe-csatornák és a vékonybél epitheliuma soha nem sérül meg magas koncentrációjuk következtében. Ez elsődlegesen más lipidek jelenlétével (foszfolipidek az epeben és a bélben lévő zsírsavakban) kapcsolódik, amelyek csökkentik az epesavak monometrikus koncentrációját, másrészt glikolipidek és koleszterin jelenlétében az epiteliális sejtek apikális membránjában, ami növeli az epiteliális sejtek rezisztenciáját a károsító az epesav-anionok hatásait.
A cholic, chenodeoxycholic és deoxycholic savak felszívódnak és enterohepatikus keringésbe kerülnek naponta 6-10 alkalommal. A litokolsav kevésbé felszívódik, és az epe mennyisége kicsi. Az epesavak összege általában körülbelül 2,5 g, és a primer epesavak napi, chol és chenodeoxycholic termelése átlagosan 330 és 280 mg.
Az epe kialakulásának szabályozása
Mint ismeretes, az epe képződés folyamatos, de ennek a folyamatnak az intenzitása változik. Fokozza az epe képződését és az ételt. A gyomor-bélrendszerben (GIT), a belső szervekben és a kondicionált reflexhatásokban a receptorok stimulálása során reflex hatása van a choleresisre.
A vegetatív szabályozást a paraszimpatikus kolinerg (az epe képződésének növelése) és a szimpatikus adrenerg idegszálak biztosítják (csökkenti az epe képződését).
A humorális szabályozást az epe önmagában végzi az epesavak enterohepatikus keringése és a negatív visszacsatolás mechanizmusa miatt (lásd fent). A Secretin serkenti az epe szekrécióját, a víz és az elektrolitok kiválasztását. A glükagon, a gasztrin és a kolecisztokinin gyengébb stimuláló hatású.
Az epehólyag kiválasztása
Az epehólyag-kiválasztás sajátos módja a molekulák és ionok eltávolításának a szervezetből, amely nem válik ki a vesén keresztül. Ezek közül a legfontosabb a koleszterin (mint önmagában és epesavak formájában) és a bilirubin, valamint a réz, a vas stb.
Az epe kiválasztásának fő összetevői
koleszterina vízben gyakorlatilag nem oldódó keveréket a vegyes micellák összetételében szállítják, amelyet a fentiek szerint, epesavakkal, foszfolipidekkel és koleszterinnel képeznek.
A foszfolipid molekulák két fontos funkciót töltenek be. Először is jelentősen növelik a koleszterin micelláris oldhatóságát, mivel a foszfolipideket tartalmazó vegyes micellák sokkal több koleszterint szolubilizálnak, mint a csak epesavmolekulákat tartalmazó egyszerű micellák. Másodszor, az epe-foszfolipidek jelenléte csökkenti a micelizáció kritikus koncentrációját és az epesavak monometrikus koncentrációját. Ennek eredményeként csökken a májsejtek felületaktivitása és citotoxicitása.
Az epe áramlásával a vegyes micellák belépnek a vékonybélbe, ahol a komponenseik további átalakulása eltérő. Az epesavak feloldják a lipideket, biztosítva abszorpciójukat, és maguk is felszívódnak a bél disztálisabb részeibe. A vízben oldhatatlan epe-foszfolipidek, a bélben hidrolizálódnak, és nem vesznek részt az enterohepatikus keringésben. Az epesavak szabályozzák kiválasztódásukat és stimulálják a szintézist.
Mivel a koleszterin kb. 1/3-a felszívódik, 2/3-a kiválasztódik. Egy felnőttnél a koleszterin egyensúlyát a felszabadulás biztosítja (kb. 600 mg / nap) vagy epesavként (kb. 400 mg / nap). Ugyanakkor az epesavak enterohepatikus keringését a koleszterin kiválasztásának késleltetett módjának tekintjük.
Meg kell jegyezni, hogy az állatokhoz képest az embereknél nagyobb arányban megfigyelhető az epe koleszterinszintje. Ez a koleszterin epesavakká történő hibás átalakulásának, valamint az epesavak szekréciójának viszonylag alacsony arányának tekinthető. Ezért az idősek kb. 25% -ában az epe túltelített a koleszterinnel, és a populáció 10-15% -ában koleszterin kövek (cholelithiasis) alakulnak ki.
Amint fentebb már említettük, az epesavak koleszterinből történő szintézisét egy negatív visszacsatolási mechanizmus szabályozza.
Az epe a bilirubin szervezetből történő eltávolításának fő módja. bilirubin - A hém lebomlásának végső terméke kémiai szerkezetében tetrapirol. A legnagyobb mennyiségű bilirubin (80-85%) az öregedő vörösvérsejtek lebomló hemoglobinjából és a csontvelőben vagy a keringési ágyban lévő újonnan kialakult vörösvértestek korai összeomlásából származik (az úgynevezett hatástalan eritropoiesis). A bilirubin fennmaradó része más hem-tartalmú fehérjék (például citokróm P-450 stb.) Pusztulásának eredményeként alakul ki a májban és sokkal kevésbé az extrahepatikus szövetekben. Meg kell jegyezni, hogy az eritrocita hemoglobin-katabolizmus a bilirubinba elsősorban a lép, a máj és a csontvelő makrofágjaiban fordul elő.
A bilirubin képződésének forrásai (százalékban)

Mint ismeretes, a nem konjugált bilirubin hidrofób (vízben nem oldódik) és egy potenciálisan toxikus anyag, amely a plazmában kering az albuminnal szorosan összefüggő állapotban és nem válik ki a vizelettel. A szervezetnek a bilirubin eliminálására való képessége a májsejtből a vérplazmából történő eltávolítással, a glukuronsavval történő konjugálással és egy már vízoldható vegyület (kötött vagy közvetlen bilirubin) felszabadításával az epe-hez kapcsolódik. A konjugációs folyamat a mikroszomális uridin-difoszfát-glükuronil-transzferázon keresztül megy végbe. A bilirubin és a glükuronsav konjugációja a legfontosabb fiziológiai jelentőséggel bír, azonban a bilirubin szulfátokkal, glükózzal és xilózzal történő konjugációja kis mennyiségben történik.
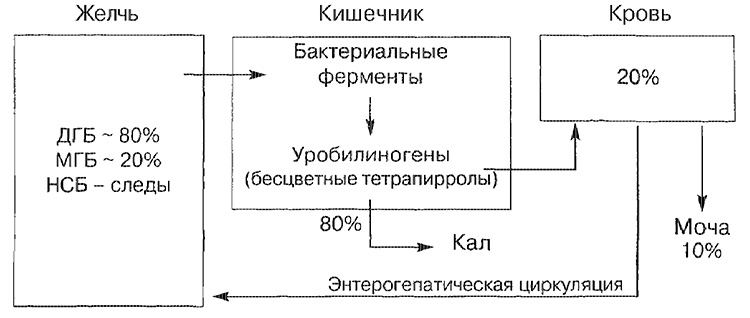
A bilirubin fő szállítási útvonalai
NSB - nem kötött bilirubin; MGB - bilirubin monoglukuronid; DGB - diglukuronid bilirubin.
A konjugált bilirubin szekréciója az epe tubulusokban az ATP-függő multispecifikus transzportfehérjék családjának részvételével történik a szerves anionokhoz.
A legtöbb bilirubin bilirubin (kb. 80%) diglucuronid bilirubin, kisebb rész formájában van jelen monoglukuronid formájában, és csak egy kis mennyiséget képvisel a nem kötött forma. A bélbe belépő bilirubin a vékonybél végső részében és a vastagbélben bakteriális enzimek (béta-glükuronidáz) révén bomlik le, amely színtelen tetrapirrol (urobilinogén) alakul át. A kialakult urobilinogének körülbelül 20% -át reszorbeálják és szükségszerűen kiválasztják a vizelettel, valamint az epe (enterohepatikus kör a keringésben).
A legtöbb szerző egyetért abban, hogy a Van Der Bergh diazo reakcióval meghatározott normál szérum teljes bilirubin-koncentráció általában nem haladja meg az 1 mg% -ot (0,3–1 mg%, vagy 5–17 µmol / l). Csak a bilirubin kevesebb, mint 5% -a van kötve. A vérben a bilirubin szintjének emelkedése (hiperbilirubinémia) és a szövetekben való felhalmozódása a sárgaság megjelenéséhez vezet, ami általában 2,5-3 mg-nál nagyobb értékekkel észlelhető.
Amint azt a fentiekben említettük, a bilirubin a vérben fehérje-kötött állapotban kering, és alig hatol be az alacsony fehérjetartalmú szövetfolyadékokra. Ebben a tekintetben a váladékok sárgabb színűek, mint a transzudátok. A bilirubin jól kötődik a rugalmas szövethez, ami magyarázza a sklerák, a bőr és az érfalak korai sárga festését hyperbilirubinemia esetén. A tartós kolesztázis zöldes bőrszín megjelenéséhez vezet, amit a biliverdin lerakódása magyaráz.
Helyénvaló hangsúlyozni, hogy az epe-csatornákban az epe-szekréció által okozott nyomás általában 150-200 mm víz. Art. 350 mm-es vízig terjed. Art. gátolja az epe szekrécióját, ami a sárgaság kialakulásához vezet. A bilirubin és az epesavak szekréciójának teljes megszűnése esetén az epe elszíneződik (az úgynevezett fehér epe).
Ezenkívül az epe a kiválasztás egyik módja növényi zsírok, lipofil gyógyszerek és metabolitjaik, különböző xenobiotikumok, amelyek a növényekben, lipofil metabolitokban, zsírban oldódó vitaminokban és szteroid hormonokban vannak jelen.
Vas- és rézegyensúly a testet a fémek epeális kiválasztása is támogatja. Mindkét kationot az ATP-vel stimulált canalicularis szivattyú választja ki az epébe. Az epével és más fémek felszabadulásával.
Epe kiválasztása
Az epe rendszerben az epe áramát a különböző szakaszokban és a nyombélben, a záróizomban, a gasztrointesztinális traktus sima izomrostjainak összehúzódásában és a csatornákban fellépő nyomáskülönbség okozza (az első egyedi izomrostok a májkapuk közvetlen közelében lévő intrahepatikus epevezetékekben jelennek meg). A teljes rendszer aktivitása általában jól koordinált és az idegrendszeri és humorális mechanizmusok irányítása alatt áll.
Az epekövekben az epét a lipidek, epesók, epe-pigmentek, stb. Koncentrációjának 10-szeres növekedésével koncentrálódik és koncentrációban növekszik. A megfelelő pillanatban az OZhP-be és tovább a duodenumba kerül. Az epe-koordináták zsírsugárzó készülékének áthaladása. A duodenum emésztési folyamatán kívül az injekciós üveg záróizomja zárva van. Ekkor az RI és a cisztás csatorna ellazult, ami lehetővé teszi a máj által termelt epe belépését az RI lumenébe. A ZH nyálkahártya elnyeli a vizet, ionokat. Ugyanakkor az epe koncentrálódik. A nyálka termékei lehetővé teszik, hogy az epe kolloid állapotban legyen.
A nyugalmi nyugalmi nyugalomban a nyomás sokkal kisebb, mint az epevezetékeknél, és 60-185 mm víz. Art. A nyomáskülönbség az epehólyagban az epe áramlásának fiziológiai alapja a zárt Oddi sphincterrel. Az emésztés során a GF csökkenése miatt a nyomás 150-260 mm-re emelkedik. st, biztosítva az epe áramlását a nyombélbe a nyugodt sphincter ampullákon keresztül. Amikor az epe elkezd áramlani a nyombélbe, a csatornákban a nyomás fokozatosan csökken (minden étkezésnél a zsírtartalom 1-2-szer csökken).
A biliáris rendszer elsődleges reakciója az élelmiszer típusától, illatától és fogadásától függően körülbelül 7-10 percig tart. Ezután jön az evakuálási időszak (a ZHP kiürítésének fő vagy a periódusa), amelynek során a ZH összehúzódásának és relaxációjának váltakozása, az epehólyag és a máj epehólyagja a duodenumba áramlik.
A cholekinesis reflexstimulációja (feltételesen és feltétel nélkül reflex) a szájüregben, a gyomorban és a nyombélben található receptorok miatt történik a hüvelyi idegeken keresztül. A humorális szabályozást főként kolecisztokinin hatására végezzük, amely stimuláló hatást fejt ki a GF-re, csökkentve ezzel a redukciót. Ezenkívül a gyomor GI gyenge vágása a gasztrin, a szekretin és a Bombezin hatására történik. Éppen ellenkezőleg, a GH, a glükagon, a kalcitonin, az antikolecisztokinin, a vasointestinalis peptid és a hasnyálmirigy polipeptid összehúzódása gátol.
A tojássárgája, a zsírok, a tej és a hús erősen stimulálják az epe kiválasztását.
A sphincter készülék és a gonadális mozgás zavarai megváltoztathatják az epe áramlásának irányát és sebességét, dyskinesiát, a hasnyálmirigy-szekréció refluxját és az AKCS duodenum tartalmát, és az epét a hasnyálmirigy csatornában, amely számos patológiai állapot és betegség kialakulásának alapja.
Az epe sókat, a konjugált bilirubint, a koleszterint, a foszfolipideket, a fehérjéket, az elektrolitokat és a vizet a hepatociták szekretálják az epehólyagba. Az epe kiválasztó készülék transzportfehérjéket tartalmaz cső alakú membrán, intracelluláris organellákés struktúrák citoszkeleton. Szoros érintkezőka hepatociták között a tubulusok lumenje elkülönül a máj keringési rendszerétől.
A cső alakú membrán az epesavak, a bilirubin, a kationok és az anionok transzportfehérjeit tartalmazza. A Microvilli növeli területét. A organellákat a Golgi készülék és a lizoszómák képviselik. A hólyagok segítségével fehérjéket (pl. IgA) szállítunk a sinusoid membránról a canalicular membránra, a sejtben szintetizált transzportfehérjék koleszterin, foszfolipidek és esetleg epesavak mikroszómákból a cső alakú membránba szállítását.
A tubulus körüli hepatocita citoplazma a citoszkeleton szerkezetét tartalmazza: mikrotubulusok, mikroszálakés közbenső szálak.
A mikrotubulusokat tubulin polimerizációjával alakítják ki, és a sejten belül egy hálózatot képeznek, különösen a bazolaterális membrán és a Golgi készülék közelében, amelyek részt vesznek a receptor-közvetített vezikuláris transzportban, a lipidek szekréciójában és bizonyos körülmények között - epesavakban. A mikrotubulus kialakulását gátolja a kolchicin.
A kölcsönhatásban lévő polimerizált (F) és szabad (G) aktinok részt vesznek a mikroszálak kialakításában. A cső alakú membrán körül koncentráló mikroszálak meghatározzák a tubulusok összehúzódását és mozgékonyságát. Falloidin, amely fokozza az aktin polimerizációját és a citokalasint B, amely gyengíti, gátolja a tubulusok mozgékonyságát és kolesztázt okoz.
A közbenső szálak citokeratinból állnak, és egy hálózatot alkotnak a citoszkeleton plazmamembránjai, magja, intracelluláris organellái és más szerkezetei között. A közbenső szálak törése az intracelluláris transzportfolyamatok megszakadásához és a tubulusok lumenének megzavarásához vezet.
A víz és az elektrolitok befolyásolják a tubuláris szekréció összetételét, áthatolva a hepatociták közötti szoros érintkezésnek a csőszerű lumen és a Disse terek közötti ozmotikus gradiens miatt. (paracelluláris áram).A szoros érintkezések integritása függ a ZO-1 fehérje plazmamembránjának belső felületéről, amelynek molekulatömege 225 kDa. A szoros érintkezések szakadásával együtt az oldott nagyobb molekulák bekerülnek a kanálba, ami az ozmotikus gradiens elvesztéséhez és a kolesztázis kialakulásához vezet. Ugyanakkor megfigyelhető a szinuszos canalicularis epe regurgitációja.
A biliáris canaliculi a csatornákba áramlik, néha kolangiolok vagy Goering csatornák. A csövek elsősorban a portálterületeken helyezkednek el, és az interlobuláris epe-csatornákba áramlanak, amelyek a máj artériájának és portálvénájának ágai kísérik az epeutak első részét, és a portál-triadok összetételében találhatók. Az interlobuláris csatornák, amelyek összekapcsolódnak, szeptális csatornákat képeznek, amíg két fő májcsatorna keletkezik, amelyek a jobb és bal lebenyből a máj porszórásában keletkeznek.
Epe szekréció
Az epe kialakulása számos illékony szállítási folyamat részvételével történik. A szekréció viszonylag független a perfúziós nyomástól. Az emberben az epe összárama körülbelül 600 ml / nap. A hepatociták az epe két frakciójának szekrécióját biztosítják: az epesavaktól függ ("225 ml / nap"), és nem függenek tőlük ("225 ml / nap"). A fennmaradó 150 ml-t naponta az epevezeték-sejtek szekretálják.
Az epesavak sóinak szekréciója az epe képződésének legfontosabb tényezője (az epesavaktól függő frakció).A víz az epesavak ozmotikusan aktív sói nyomán mozog. Az ozmotikus aktivitás változásai szabályozzák a víz áramlását az epébe. Egyértelmű összefüggés van az epesavak sóinak szekréciója és az epe áramlása között.
Az epesavaktól független epefrakció meglétét igazolja az epe képződésének lehetősége, amely nem tartalmaz epesavak sóit. Így az epesavak sóinak kiválasztódása ellenére is lehetséges az epe áramlásának folytatása; a víz kiválasztása ebben az esetben más ozmotikusan aktív oldható anyagok, például glutation és bikarbonát miatt van.
Az epe kiválasztásának sejtes mechanizmusai
A hepatocita egy poláris szekréciós epiteliális sejt, amely bazolaterális (szinuszos és laterális) és apikális (cső alakú) membránokat tartalmaz.
Az epe képződése magában foglalja az epesavak és más szerves és szervetlen ionok befogását, a bazolaterális (szinuszos) membránon, a citoplazmán és a tubuláris membránon keresztül történő szállítását. Ezt a folyamatot a hepatocita és a paracelluláris térben lévő víz ozmotikus szűrése kíséri. A szinuszos és cső alakú membránok transzportfehérjeinek azonosítása és jellemzése összetett. Különösen nehéz a tubulusok szekréciós készülékének vizsgálata, azonban a mai napig számos tanulmányban kifejlesztették és bizonyítottan megbízhatónak bizonyult egy rövid életű kultúrában megduplázott hepatociták megszerzésére szolgáló technika.
Az epe képződésének folyamata attól függ, hogy "bizonyos hordozófehérjék jelen vannak a bazolaterális és tubuláris membránokban. Na +, K a szekréció hajtóerejének szerepe." + - A bazolaterális membrán ATPáza, amely kémiai gradienst és potenciális különbséget biztosít a hepatocita és a környező tér között. Na +, K + - Az ATPáz három intracelluláris nátriumiont cserél két extracelluláris káliumionra, fenntartva a nátrium (magas külső, alacsony belső) és kálium (alacsony külső, magas belsejében) koncentrációs gradiensét. Ennek eredményeképpen a sejttartalom negatív töltéssel rendelkezik (–35 mV) az extracelluláris térhez képest, ami megkönnyíti a pozitív töltésű ionok rögzítését és a negatív töltésű ionok kiválasztását. Na +, K + -ATPáz nem detektálható a cső alakú membránban. A membrán áramlása befolyásolhatja az enzimaktivitást.
Rögzítsen egy szinuszos membrán felületén
A bazolaterális (szinuszos) membrán számos transzportrendszerrel rendelkezik a szerves anionok rögzítésére, amelyek szubsztrátspecifitása részben átfed. A hordozófehérjék jellemzését korábban az állati sejtek tanulmányozása alapján adtuk meg. A humán szállítási fehérjék közelmúltbeli klónozása lehetővé tette a funkciók jobb jellemzését. A szerves anionok transzportfehérje (szerves anion szállító fehérje - OATP) nátriumfüggetlen, számos vegyület molekuláját hordozza, beleértve az epesavakat, bróm-szulfaleint és valószínűleg bilirubint. Úgy véljük, hogy a bilirubin transzportját a hepatocitába más hordozók is végzik. A taurinnal (vagy glicinnel) konjugált epesavakat a nátrium / taurokolát transzportfehérje (NTCP nátrium-epesav-transzport fehérje) segítségével végezzük.
Az ionátvitel a basolaterális membránon keresztül olyan fehérjét tartalmaz, amely kicseréli a Na + / H + -ot és szabályozza a sejten belüli pH-t. Ezt a funkciót a Na + / HCO3 - transzport fehérje is elvégzi. A bazolaterális membrán felszínén szulfátok, nem észterezett zsírsavak és szerves kationok is rögzíthetők.
Intracelluláris szállítás
Az epesavaknak a hepatocitába történő szállítását citoszol fehérjékkel végzik, amelyek közül a fő szerepet a Z-hidroxiszteroid dehidrogenáz adja. A glutation-S-transzferáz és a zsírsavkötő fehérjék kevésbé fontosak. Az endoplazmatikus retikulum és a Golgi készülék részt vesz az epesavak transzferében. Nyilvánvaló, hogy a vezikuláris transzport csak akkor aktiválódik, ha az epesavak jelentősen injektálódnak a sejtbe (fiziológiai értéket meghaladó koncentrációban).
A fehérjék folyadékfázisban és ligandumokban, például IgA-ban és kis sűrűségű lipoproteinekben történő szállítását vezikuláris transzcitózissal hajtjuk végre. A bazolaterális és a cső alakú membrán közötti átviteli idő körülbelül 10 perc. Ez a mechanizmus az epe teljes áramának csak egy kis részét képezi, és a mikrotubulusok állapotától függ.
Tubuláris szekréció
A cső alakú membrán a hepatocita plazmamembrán speciális szakasza, amely olyan transzportfehérjéket (többnyire ATP-függő) tartalmaz, amelyek felelősek a molekulák epe-be történő átadásáért egy koncentrációs gradienssel szemben. Az enzimek, mint például az alkalikus foszfatáz, a GGTP is a lokális membránban találhatók. A glükuronidok és glutation-S-konjugátumok (például bilirubin-diglukuronid) transzferét egy szerves anionok (cMOAT-mirigy) Canalicular multispecifikus transzportfehérje segítségével végezzük; transzporter - SWAT), amelynek funkcióját részben negatív intracelluláris potenciál szabályozza. Az epesavaktól független epe áramát a glutaion transzportja, valamint a bikarbonát canalicularis szekréciója határozza meg, esetleg a Cl - / HCO 3 - cseréjével.
A P-glikoprotein család két enziméhez fontos szerepet játszik az anyagoknak a tubuláris membránon keresztül történő szállítása; mindkét enzim ATP-függő. A multidrug rezisztencia fehérje 1 (MDR1) multidrog rezisztencia fehérje szerves kationokat transzferál, és a citosztatikus gyógyszereket is eltávolítja a ráksejtekből, ami a kemoterápiával szembeni rezisztenciát (így a névfehérjét) okoz. Az MDR1 endogén szubsztrátja nem ismert. Az MDR3 foszfolipideket szállít és foszfátként működik a foszfatidil-kolin számára. Az MDR3 funkcióját és annak fontosságát az epe-ben lévő foszfolipidek szekréciójában az egereken végzett kísérletekben, amelyekben nincs mdr2-P-glikoprotein (humán MDR3 analóg). Az epeben lévő foszfolipidek hiányában az epesavak károsítják az epe epitéliumot, a ductulák gyulladását és a periductularis fibrózist.
A víz és a szervetlen ionok (különösen a nátrium) az ozmotikus gradiens mentén a biliáris kapillárisokba kerülnek, a negatív töltésű, féligáteresztő, szoros érintkezőkön keresztül történő diffúzió útján.
Az epe szekréciót számos hormon és másodlagos hírvivő szabályozza, beleértve a cAMP-t és a protein-kinázt. Az epe áthaladása a tubulusokon a mikroszálaknak köszönhető, amelyek a tubulusok mozgékonyságát és összehúzódását biztosítják.
Ductile szekréció
A távoli csatornák epithelialis sejtjei bikarbonátban gazdag titkot képeznek, amely módosítja a cső alakú epe összetételét (az úgynevezett ductular áram, epe).A szekréciós eljárás cAMP-t, néhány membránszállító proteint termel, beleértve a Cl - / HCO 3 -, és a fehérjét is a cisztás fibrózisban a transzmembrán vezetés szabályozója -membráncsatorna Cl-hez, állítható cAMP. A szekretin stimulálja a ductularis szekréciót.
Feltételezzük, hogy az ursodeoxikolsav aktívan abszorbeálódik a duktuláris sejtekben, kicserélve bikarbonátokra, újrahasznosul a májban, majd ismét kiürül az epébe ("cholehepatikus shunt"). Talán ez magyarázza az ursodeoxikolsav koleretikus hatását, melyet kísérleti cirrhosisban magas biliumbikarbonát-szekréció kísér.
Az epe-csatornákban az epe kiválasztását okozó nyomás általában 15-25 cm víz. Art. Növelje a nyomást 35 cm-es vízre. Art. az epe szekréció szuppressziójához, sárgaság kialakulásához vezet. A bilirubin és az epesavak szekréciója teljesen leállítható, míg az epe színtelen lesz (fehér gallér)és hasonlít a nyálkafolyadékra.
Az epe egy izoszmotikus plazmafolyadék, amely vízből, elektrolitokból és szerves anyagokból (epesavak, foszfolipidek, koleszterin, bilirubin) áll. Az epesavak (vagy ezek sói) az epe fő szerves összetevői. Az epesavak két forrásból származnak: (1) a máj koleszterinéből szintetizált primer epesavak (chol és chenodeoxycholic); (2) a másodlagos epesavak (deoxikolsav, lithocholic és ursodeoxycholic) a bélbaktériumok elsődleges epesavak hatására alakulnak ki. Maguk az epesavak két fontos komponensből állnak, amelyek meghatározzák fiziológiai és fizikai-kémiai tulajdonságaikat: (1) egy szteroid mag hidroxil szubsztituensekkel; (2) egy alifás oldallánc (7-4. Ábra).
Ábra. 7-4.
Az epesavak két komponensből állnak - egy hidroxil-végű magból és egy alifás oldalláncból. Az ábrán a kolinsavat a 3-hidroxi-sav (3'-, 7p-, 12p-OH) példaként mutatjuk be. További példaként említhetők az epesavak, amelyek deoxikolátot (3p, 12a-OH), chenodeoxi-kolátot (3p-, 7p-OH) és lithocholate (3p-OH) tartalmaznak.
A legtöbb emlősben a primer epesavak három-hét hidroxil-szubsztituenst tartalmaznak, amelyek száma befolyásolja vízoldhatóságukat (hidrofilitás). Nem sokkal a kialakulás után a primer epesavak módosulnak a terminális karboxilcsoportban. Ez a másodlagos epesavak enterohepatikus keringésének májfázisában és glicinnel vagy taurinnal való konjugáció során következik be. Az alifás oldallánc hidrofil (hidroxil komponensei és amidkötése) és hidrofób (szteroid mag) komponensek jelenléte lehetővé teszi a konjugált epesav molekulák amfoter vegyületként való működését. Ez lehetőséget ad számukra a micellák (polimolekuláris aggregátumok) kialakítására a kritikus micelláris koncentráció felett. Az epesavak molekulái viszont képesek más amfoter anyagok (koleszterin, foszfolipidek) feloldására vegyes micellák képződésével. Az epesavak ilyen detergensszerű szerepe fontos az epe fizikai-kémiai állapotának stabilizálásához, az emésztéshez és a zsírok felszívódásához.
A koleszterinből származó epesavak szintézisét egy negatív visszacsatolási mechanizmus szabályozza, bár a molekuláris és biokémiai szintek szabályozásának jellege még nem teljesen ismert. A koleszterin mikroszomális 7a-hidroxilezése kritikus lépés az epesavak szintézisében. Az epehólyagkövek feloldására használt chenodesoxycholic acid gátolja az epesavak szintézisét, és ezáltal növeli a vér koleszterinszintjét. Az ursodeoxikolsav használatakor az ilyen változásokat nem tartják fenn hosszú távú kezelés esetén sem.
Az epe képződése a hepatocita membrán szinuszos és tubuláris felületén történik, és egyaránt intracelluláris és paracelluláris folyamat. A vesékben a glomeruláris szűréssel ellentétben, amely hidrosztatikus erők hatására passzívan, az epe képződése során a szerves és szervetlen komponensek aktív átvitele történik a tubulusok lumenébe és a passzív vízszállításba. Így az epe kiválasztódási folyamatai hasonlítanak a hasnyálmirigy acini szekréciójához, a vese-tubulusok epitéliumához. A cső alakú epe képződése két típusra osztható (7-5. Ábra): (1) az epesavak szekréciójától függően az epe képződése, amely a tubulusokban szekretált epe mennyiségének és a szekretált epesók mennyiségének aránya; (2) az epesav-szekréciótól függetlenül az epe képződése, amely a szervetlen elektrolitok és más anyagok aktív szekréciójaként jeleníthető meg, és ez a vonal y-metszéspontjaként jelenik meg. Más szavakkal, a savas szekrécióval összefüggő epe képződés az epe áramlási sebessége, attól függően, hogy az epe-csatornákban az ozmotikusan aktív epesók jelen vannak-e, és epe-sók hiányában a savszekrécióval nem összefüggő epe képződés. Az epe képződésének aránya és az epesavak sóinak képződése nem-lineáris kis mennyiségű szekretált epével és nem felel meg az 1. ábrán látható lineáris kapcsolatnak. 7-5. Ezért az epe képződésének mindkét típusát kölcsönösen összefüggő indikátoroknak kell tekinteni.
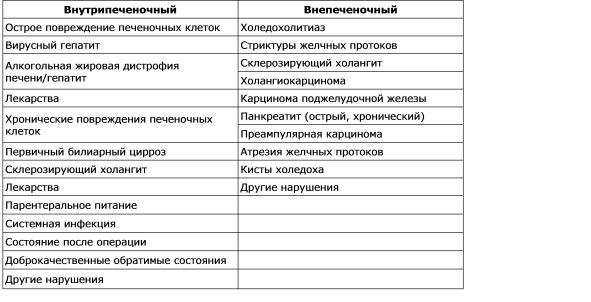
Az epe képződésének megsértését cholestasis-nak hívják. A kolesztázis kialakuló patológiás, fiziológiai és klinikai megnyilvánulásának sorrendje az oka. A pericentrális hepatociták tubulusaiban kimutatható a hepatobiopszia epe anyagának morfológiai vizsgálata, a tubulusok dilatációja figyelhető meg, és az ultrastruktúra vizsgálata a mikrovillák számának csökkenését mutatja. A kolesztázis a hepatocita (intrahepatikus kolesztázis) szintjén az epe képződésének funkcionális hibája, valamint az epe (extrahepatikus kolesztázis) szerves vagy mechanikai károsodott szekréciója és kiáramlása. Az intrahepatikus és extrahepatikus kolesztázis leggyakoribb okai a 2. táblázatban találhatók. 7-2. Számos mechanizmus van, amelyek fontos szerepet játszanak az intrahepatikus kolesztázis patogenezisében: a szinuszos membrán működésének károsodása és károsodása; a hepatocita intracelluláris organellák funkcióinak megszakítása; a cső alakú membrán károsodása és megszakadása. Tehát nem létezik egyetlen mechanizmus a kolesztázis különböző klinikai helyzetekben, és a mechanizmusok sokasága különböző rendellenességekhez vezethet. Klinikailag a kolesztázist számos anyag, köztük a bilirubin, az epesók, a koleszterin szintjének emelkedése jellemzi. A vér cholestázissal végzett biokémiai analízisében az alkalikus foszfatáz aktivitás és a bilirubin koncentráció aránytalan növekedése figyelhető meg az aminotranszferázok szintjének változásával párhuzamosan, amit az alábbiakban tárgyalunk.
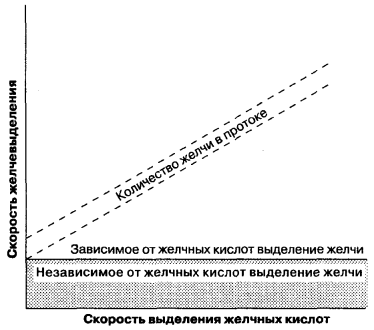
Ábra. 7-5.
Az epesav-szekrécióval összefüggő epe képződés, az epesav-szekréciótól független epe képződés. (By: Moseley R. H., Bile szekréció. Yamada T., Alpers D. H., Owyang C., Powell D. W., Silverstein F. E., szerk., Textbook of Gastroenterology, 2. kiadás, Philadelphia: J. B. Lippincott, 1995: 387.)
7. - 2. táblázat.
A májban a legfontosabb emésztési gyümölcslé képződik - epe.
Az epét a hepatociták termelik a víz, koleszterin, bilirubin és kationok aktív és passzív szállításával. A koleszterinből származó hepatocitákban elsődleges epesavak képződnek - cholic és deoxycholic. A vízben oldódó komplexet bilirubinból és glükuronsavból szintetizáljuk. Belépnek az epe kapillárisokba és csatornákba, ahol az epesavak glicinnel és taurinnal kombinálódnak. Ennek eredményeként glikokolsav és taurokolsav keletkezik. A nátrium-hidrogén-karbonátot ugyanazok a mechanizmusok képezik, mint a hasnyálmirigyben.
Az epét mindig a máj által termeli. Napján körülbelül 1 liter képződik. A hepatociták kiválasztják az elsődleges vagy a máj epét. Ez a folyadék arany sárga lúgos reakció. PH = 7,4 - 8,6. 97,5% vizet és 2,5% szilárd anyagot tartalmaz. A száraz maradék tartalmaz:
1. Ásványi anyagok. Nátrium, kálium, kalcium kationok, bikarbonát, foszfát anionok, klórionok.
2. savas savak - taurokolikus és glikokolsav.
3. Epe pigmentek - bilirubin és oxidált formája biliverdin. A bilirubin epe színt ad.
4. Koleszterin és zsírsavak.
5. Karbamid, húgysav, kreatinin.
Mivel az emésztőrendszeren kívül, a közös epevezeték szájánál elhelyezkedő Oddi gömbölye zárva van, az ürülék ebből a részéből halmozódik fel. epehólyag. Itt a víz újra felszívódik belőle, és az alapvető szerves komponensek és a mucin tartalma 5-10-szeresére nő. Ezért a cisztás epe 92% vizet és 8% száraz maradékot tartalmaz. Sötétebb, vastagabb és viszkózusabb, mint a máj. Ezen koncentráció miatt a húgyhólyag 12 órára felhalmozódhat epe. Az emésztés során Oddi sphinctere és a húgyhólyag nyakán lévő Lutkens sphincterje nyitott. Az epe belép a nyombélbe.
Bile érték:
1. Az almasavakat a zsírok egy részét emulgeálják, a nagy zsírrészecskéket finom cseppekké alakítják.
2. Aktiválja a bél- és hasnyálmirigy-lé, különösen a lipáz enzimeit.
3. Az epesavakkal kombinálva a hosszú láncú zsírsavak és a zsírban oldódó vitaminok felszívódása az enterociták membránján keresztül történik.
4. Zhelchka elősegíti a trigliceridek újraszintézisét enterocitákban.
5.Inaktiválja a pepszint, és semlegesíti a gyomorból származó savanyú kémát. Ez biztosítja a gyomor és a bél emésztése közötti átmenetet.
6.Serkenti a hasnyálmirigy és a béllé szekrécióját, valamint az enterociták elszaporodását és szétválasztását.
7. Megerősíti a bélmozgást.
8. Bakteriosztatikus hatással van a bél mikroorganizmusaira, és így megakadályozza a rothadásos folyamatok kialakulását.
Az epe képződésének és az epe kiválasztásának szabályozását főleg humorális mechanizmusok végzik, bár az idegesek bizonyos szerepet játszanak. A májban az epe képződésének legerősebb serkentője az epesavak, amelyek a bélből a vérbe szívódnak. A szekretin fokozza azt is, amely hozzájárul az epe nátrium-hidrogén-karbonátjának növekedéséhez. A vagus ideg stimulálja az epe termelését, a szimpatikus gátlást.
Amikor a chyme belép a nyombélbe, az I-sejtek elkezdik felszabadítani a kolecystokinin-pancreozymin i-sejteket. Különösen ezt a folyamatot serkenti zsírok, tojássárgája és magnézium-szulfát. A CCK-PZ erősíti a húgyhólyag simaizomainak összehúzódását, az epevezetékeket, de ellazítja Lutkens és Oddi sphinctereit. Az epe a bélbe kerül. A reflex mechanizmusok kis szerepet játszanak. A Chyme irritálja a vékonybél kemoreceptorjait. Az impulzusok belépnek a medulla emésztőközpontjába. Tőle a hüvelyben vannak az epeutak felé. A sphincters pihen, és a húgyhólyag-szerződés sima izmait. Ez elősegíti az epe kiválasztását.
A legsúlyosabb betegségek a hepatitis és a cirrózis. A hepatitisz leggyakrabban a fertőzés (A, B, C fertőző hepatitis) és toxikus termékek (alkohol) hatásának következménye. A hepatitisben a hepatociták érintettek, és minden májfunkció romlik. A cirrózis a hepatitis következménye. Az epehólyag-kiválasztás leggyakoribb megsértése a cholelithiasis. Az epekövek nagy részét koleszterin képezi, mivel az ilyen betegek epe túltelített.
A máj a szervezet legnagyobb szerve és központi szerepet játszik az anyagcserében. Számos funkciót tölt be, részt vesz a fehérjék, szénhidrátok, zsírok, hormonok és vitaminok metabolizmusában, valamint számos endogén és exogén anyag semlegesítésében. Ezeket a folyamatokat a fiziológiai kémiai tankönyvekben és ebben a szakaszban ismertetik emésztőrendszer csak figyelembe fogjuk venni kiválasztási májfunkcióazaz epeválasztás. epe vízből, ásványi sókból, nyálkából, koleszterin lipidekből és lecitinből és két fajta összetevőből áll - epesavak és bilirubin pigment. Az epesavak detergensek, emulgeáló hatásuk pedig fontos szerepet játszik a lipid emésztésben. A bilirubin a szervezetből eltávolítandó hemoglobin lebontásának végterméke.
^
Epe kialakulása
Funkcionális anatómia.Májsejtek (Hepatociták)egy cellát képeznek egy cellának a vastagságában, elválasztva keskeny résekkel (Disse tér)a befejezett
^ 766 VIII. RÉSZ. ÉLELMISZER, DIGÉZÉS ÉS EXTRACT
Vérrel szinuszhullámkapillárisokkal egyenértékű. A sinusoidok falaiban pórusok vannak, amelyeken keresztül az ilyen nagy makromolekulák, mint albumin és lipoproteinek áthaladhatnak. A legkisebb kanaltsam-a két szomszédos hepatociták plazmamembránja által határolt epe-kapillárisok - az epe nagyobbak goring's canaliculi,a falak, mint a nagyobbak interollege tubulusés epevezetékekköbös szekréciós sejtek alkotják. A kisméretű tubulusok a máj lebenyein belül és közöttük nagyobbak, egyesülnek májcsatorna.Ebből a csatornából indul cisztás csatornaaz epehólyaghoz. Az egyesülés után a máj- és a cisztásvezetékek formája közös epevezetéknyílás a nyombélben a Vater papillájának tetején, a hasnyálmirigy-csatorna mögött vagy annak közelében (29.1. ábra).
^ Az epe funkciói.A Bile számos fontos funkciót hajt végre. Vele a végtermékek megjelenneka bilirubin, valamint a kábítószerek és toxinok. kiosztás aepe koleszterinfontos szerepet játszik egyensúlyának szabályozásában. Epesavaknélkülözhetetlen az emulgeáláshoz és a zsírfelszíváshoz. Ezenkívül az epe vizet, ásványi sókat és nyálkákat tartalmaz. A nap 24 órájában 600 mlaz epe, és ennek az összegnek 2/3-a a tubulusokból, a nagyobb csatornák 1/3-a.
^ Tubularis epe megközelítőleg azonos mennyiségben képződik két különböző mechanizmus részvételével, az epefüggő és az epefüggő (29.27. ábra).
^ Epesavfüggő szekréció. Közvetlen kapcsolat áll fenn az epe áramlási sebességeés epesavszekréció.A canalicularis epe esetében az epesavak koncentrációja 100-szor nagyobb, mint a portálvérben, ezért ezeket úgy tekintik, hogy a szekretált aktív transzport vektorral.Az osmotikus gradiens mentén az epesavakat követve a víz a csövekbe rohan, így az epe izotóniás a vérrel.
Az epesavaknak két forrása van. Először is a hepatocitákban a koleszterinből de novo-t szintetizálják magukban a 7-hidroxiláz részvételével. Ez az enzim kulcsszerepet játszik és szabályozza az epesavak szintézisének sebességét egy visszacsatolási mechanizmuson keresztül. Másodszor, a hepatociták képesek aktívan felszívni az epesavakat portál vérés kivesszük azokat tubulusokba (lásd még a 29.29. ábrát). Ez az extrakció nagyon hatékony; a vér egyetlen áthaladásával, az epesavak 80% -a kivonódik belőle. ezért
Az epesavak koncentrációja a perifériás vérben sokkal alacsonyabb, mint a portálrendszerben. Mivel az epesavak a vérből 6-szor gyorsabban extrahálódnak, mint a tubulusokba, az utóbbi eljárás korlátozza az epesavszekréció sebességét.
^ Az epesavaktól független szekréció. az
Ez az eljárás Na +, Cl-, HCO3- és vízionokat tartalmaz. A hajtóerő aktív szállítás Να + , esetleg bikarbonáttal. Az epesav-független szekréció különösen \\ t secretin.
Az epesavak mellett a bilirubin, a koleszterin és a foszfolipidek (főként lecitin) aktívan szekretálódnak a canaliculi-ba (29.27. Ábra). Vízben nem oldódik ("Közvetett") bilirubin,amelyek többsége az idősebb vörösvértestek hemoglobinjából képződik, belép a hepatocitákba az albuminnal társult kolloid aggregátum formájában. Napi képződése körülbelül 4 g / testtömeg kg vagy 200-300 mg / nap. A hepatocitákban a bilirubin 80% -a konjugált. aglukuronsav és kis mennyiségű kénsavval. Ilyen konjugált
^ 29. FEJEZET. A GASTROINTESTINAL TRACT 767 FUNKCIÓI
A bilirubin kiválasztódik az epével. ("Közvetlen" bilirubin).Általában a gyógyszereket és a toxinokat ugyanúgy eltávolítják.
^ Az epevezeték epe módosítása
(29.27. ábra). A csatornákban, ahol a tubulusok nyitva vannak, az elsődleges epe módosul. Ez a folyamat hasonlít a glomeruláris szűrlet módosítására a vese-tubulusokban (785. o.), És hasonló módon kiszámítja azt vámkezelésaz inulin helyett csak epe esetén inert anyagot használnak eritritvagy mannit,amelyek a tubulákba szekretálódnak, de nem reagálnak. Hasonló tanulmányok kimutatták, hogy az NSO ^ aktív szekréciójával a csatornákba kb. secretin.
^
Máj- és epehólyag-epe
A máj epe összetétele(29.3. lap). A máj által 0,4 ml / perc sebességgel választott epével aranyszínű, ami a bilirubin jelenlétében magyarázható. Az elektrolitok koncentrációja ebben az epében ugyanaz, mint a plazmában, kivéve azt a tényt, hogy kétszer annyi HCO3-ot tartalmaz, és némileg kisebb, mint a CI ~. szinte kizárólag epesavak, koleszterin és foszfolipidek.
 |
^ Epesavaka hidroxilezés és a karboxilcsoport hozzáadása következtében a májban koleszterinből képződnek. A májban képződő savak elsődleges epesavak;ezek közé tartozik chenodezoxikólsav(dioxyform) és cholic(Trioksiforma) sav.A májban nem szabad formában, hanem glicinnel és taurinnal alkotott konjugátumok formájában, és a glicinnel alkotott konjugátumok háromszor nagyobbak, mivel a taurin mennyisége korlátozott. A konjugált epesavak vízben jobban oldódnak,
Nem konjugált, és nagyobb képessége van az epe sóinak kationokkal történő disszociálására és formálására, főként Na + ionokkal. Savas környezetben (pH 4,0) az epesav-sók oldhatatlanok és kicsapódnak, de fiziológiai pH-értékeken (a vékonybélben) jól oldódnak.
Az ileum távoli részében és a vastagbélben az elsődleges epesavak néhány sója anaerob baktériumok hatására dehidroxileződik, és átalakul másodlagos epesav-lithocholic(monooxiform) és dezoxikolinsav(Dioksiforma). A chenodeoxycholic, cholic és deoxycholic savak 2: 2: 1 arányban vannak jelen. A litokolsav csak néhány frakcióban van jelen, mivel a legtöbbjük kiválasztódik.
Az epesavak emulgeáló hatása a zsírokra főként a formázási képességén alapul micellák.Az epesavmolekuláknak háromdimenziós szerkezete van, amelyben a hidrofil karboxil- és hidroxilcsoportok a molekula ugyanazon oldalán vannak, és a molekula hidrofób része (szteroid mag, metilcsoportok) az ellenkező oldalon van, ami miatt az epesavmolekulák rendelkeznek és hidrofil,és lipofil tulajdonságok.Ennek a szerkezetnek köszönhetően az epesavak molekulái detergensként működnek: a lipid és a vizes fázisok felületén szinte monomolekuláris filmet képeznek, amelyben a hidrofil csoportok a vizes és a lipofil lipid fázishoz fordulnak. A vizes fázisban az epesavak rendezett aggregátumokat képeznek. -mitsellyfeltéve, hogy azok koncentrációja eléri a bizonyos szintet, amit hívnak kritikus micella koncentráció(1-2 mmol / l). A micellák belső, lipofil régiója tartalmazhat lipidek,például koleszterin és foszfolipidek; az ilyen micellákat kevertnek nevezik (29.28. ábra). Maga a koleszterin vízben nem oldódik, de a micellák összetételében oldható. Ha a koncentrációja meghaladja a micellák kapacitását, kristályos csapadékot képez; ez a folyamat a koleszterin-epekő kialakulásának alapját képezi (769. o.).
^ A cisztikus epe összetétele (29.3. lap). Az epehólyag kapacitása mindössze 50-60 ml. Ugyanakkor a máj 600 ml / nap sebességgel szekretál az epe, és ennek a mennyiségének a fele áthalad az epehólyagban, mielőtt belépne a vékonybélbe. Az epehólyagba belépő epe térfogata és kapacitása közötti különbséget kompenzálják rendkívül hatékony reabszorpcióvíz az epehólyagban. Néhány óra múlva a víz 90% -át visszavezethetjük az epéből. a
^
768 VIII. RÉSZ. ÉLELMISZER, DIGÉZÉS ÉS EXTRACT
Ez a szerves anyag az epehólyagban marad és koncentrációja az epében nő. A reabszorpció hajtóereje a nα * ionok aktív szállításaa sejtek bazális és oldalsó membránjába ágyazott „szivattyú” részvételével, és a (Na + -K +) - ATPáz által aktivált. A Na + ionok után a Cl - és HCO 3 - ionok diffundálódnak az elektromos gradiens irányában, vagy a hordozók szállítják. A HCO 3 újbóli felszívódása következtében az epehólyag pH-ja 6,5-re csökkent 8,2-re a máj epében. Az extracelluláris térben az epehólyag epitheliumban a Na + ionok magas koncentrációjának kialakulása következtében oszmotikus gradiens alakul ki, ami víz szivattyúzásához vezet, amely azután áramlik a kapillárisokba (751. oldal).
^ Motilitás epehólyag. Az éhgyomri állapotban az epe felhalmozódik az epehólyagban, és
Az étkezési időt az epehólyag összehúzódása következtében osztják szét. Az epehólyag összehúzódó aktivitásának fő ösztönzője kolecisztokinin,a duodenum nyálkahártyája által kiváltott zsírszegényt kivéve. Az epehólyag-összehúzódásokat bizonyos mértékig stimulálják. vándorló idegés paraszimpatikus.A zsíros ételek érintkezésbe kerülnek a bél nyálkahártyájával 2 percen belül, és 15-90 perc után a hólyag teljesen kiürül. Az epehólyag motilitása két folyamatot foglal magában. Először egy tonikus összehúzódás alakul ki, aminek következtében csökken az epehólyag átmérője, és ezután periodikus összehúzódások lépnek fel erre a hatásra, amelynek gyakorisága 2-6 / perc. E két folyamat eredményeként 25–30 mm Hg nyomás keletkezik. Art.
^
Enterohepatikus keringés
Epesav-keringés(29.29. ábra). Az epesavak a duodenumba válnak vegyes micellaként. Annak ellenére, hogy a gyomor tartalmával az epesavak hígulnak, a bélben a koncentráció körülbelül 10 mmol / l, és meghaladja a micellák képződésének kritikus koncentrációját. Itt a koleszterin és a lecitin mellett a micellák közé tartoznak a hidrolitikus zsírbontás termékei is. zsírsavakés monogliceridek.A micellák kezdeti érintkezésénél a bélfal
^
29. FEJEZET. A GASTROINTESTINAL TRACT 769 FUNKCIÓI
A lipidek a kefe szegélyének membránján belépnek enterocitákba, és az epesavak a béllumenben maradnak, de a belek további áthaladásával az epesavakat az aktív és passzív transzport veszi fel.
Az epesavak mintegy 50% -a passzívan szívódik fel a bélben. Az epesav-konjugátumok hasítása és az utóbbiak bélbaktériumok hatására történő dehidroxilezése eredményeként a lipidek oldhatósága nő, és a passzív diffúzió elősegül.
^ Aktív szívás az epesavak kizárólag a terminális ileumban fordulnak elő - ritka jelenség, amely csak a B 12-vitamin felszívódása miatt ismert. Aktív felszívódás csak azoknak az epe-savaknak, amelyek passzív abszorpcióját gátolják, például a taurin konjugátumok, nagy aktivitásúak. Az epesavak felszívódási folyamatát a terminális ileumban az aktív transzport jellemző jelei jellemzik: telítési kinetika és versenyképes gátlás. Kis mennyiségű epesav (7-20%) nem szerepel az aktív vagy passzív abszorpcióban, és a szervezetből eliminálódik.
Az epesavak jelenléte a vastagbélben fontos szerepet játszik a széklet konzisztenciájának szabályozásában. Amikor a dioxin-sav koncentrációja a vastagbélben nagyobb, mint 3 mmol / l, jelentős mennyiségű elektrolit és víz válik ki a bél lumenébe, ami hasmenéshez vezet. Ennek kifejezett formája "Hologennogo" hasmenésmegfigyelhető a terminális ileum reszekciójával vagy betegségével, és annak kezelésére az epesavak kötődésével kolesztiramin-ioncserélő alkalmazásával.
Lenyelve felszívódott epesavakat a májbaa konjugátumok újonnan képződnek, és néhány másodlagos epesav hidroxilezésen megy keresztül. Az epesavak veszteségét a székletben (0,2-0,6 g / nap) kompenzálja a szintézisük.
^ Az epesavak teljes összege a szervezetben körülbelül 3,0 g, ez nem elegendő az étkezés utáni lipolízis biztosításához; különösen a zsíros ételek fogyasztása 5-ször több epesavat igényel. Azonban a test nem hiányos az epesavakban, mivel sokszor keringenek a belekben és a májban. (enterohepatikus keringés).Az epesavak medencéjének teljes gyakorisága a táplálkozási módtól függ és napi 4-12 ciklusban van.
^ A bilirubin keringése. Gall pigment bilirubin,mint az epesavak és a lipidek, belép a bélbe glzuronid formájában. Csak kis mennyiségű poláris vegyület
Reagál az epehólyagba és a vékonybélbe. A terminális ileumban és (leginkább) a vastagbélben a bilirubin konjugátumot bakteriális hidrolázokkal hasítjuk. Ugyanakkor a bilirubin átalakul urobilinogénszint,amely a bilirubin egyéb bomlástermékeivel együtt barna színt ad a székletnek. Az urobilinogén kevesebb mint 20% -a felszívódik vissza, és ebből az összegből körülbelül 90% -kal visszatér a májba, és visszatér az epe-re, és a fennmaradó 10% -aa kiválasztódik a vizelettel.
Kórélettani szempontok. A vizeletben az urobilinogén szintje megemelkedhet májbetegség,a bilirubin kiválasztásának megsértése. Az urobilinogén teljes hiánya a vizeletben, a széklet és a sárgaság világos színe az epehólyag teljes elzáródása;ebben a helyzetben a bilirubin egyáltalán nem lép be a bélbe, és az urobilinogén nem képződik.
A biliáris rendszer normális fiziológiájának legismertebb és legelterjedtebb megsértése a koleszterin kicsapása koleszterin-epekő kialakulásával. A koleszterin, mint a lecitin, csak vegyes micellák részeként oldott állapotban van. Ha a koncentráció megnő xo.jecmepo.ia vagy az epesavak vagy lecitin koncentrációja a kritikus szint alatt marad,a koleszterin kicsapódik. A relatív koleszterin-tartalom növekedését okozó tényezők közé tartoznak az ösztrogének, a szénhidrát-étrend, a túlsúly és az epesavak koncentrációját csökkentő folyamatok, például az ileum gyulladása. (Crohn-betegség)vagy resection. Bizonyos esetekben az orális epesav kiegészítés elegendő lehet ahhoz, hogy a lithogén epe alitogénvé váljon, amelyben a koleszterin kövek feloldódhatnak. Ehhez a legmegfelelőbbek a chenodeoxycholic és urodesoxycholic savak, mivel nem okoznak hasmenést.
A csökkent bilirubin metabolizmus klinikai megnyilvánulása a sárgaság. A sárga bőr a bilirubin plazmaszintjének emelkedésével jár, ami a következő esetekben fordulhat elő:
fokozott bilirubin képződése a fokozott eritrocita lebomlás következtében (hemolitikus sárgaság);
a konjugációs folyamat megszakítása vagy a bilirubin hepatocitákban történő szállítása következtében, mint például sárgaság terhesvagy gilbert anyasági sárgasága;
az epehólyag késleltetett kiáramlása miatt, például epekövek vagy az epeutak régiójában lokalizált tumorok miatt (obstruktív sárgaság).