La formación de la bilis. La fisiología de la formación biliar. Formacion biliar
La formación de bilis se produce en el hígado como resultado del transporte activo y pasivo de sustancias (agua, glucosa, creatinina, electrolitos, hormonas, vitaminas, etc.) que circulan en la sangre a través de las células y los contactos célula - célula, así como la secreción activa de componentes biliares (ácidos biliares) por hepatocitos. y la reabsorción de agua y ciertas sustancias de los pequeños conductos biliares y cálculos biliares. La importancia fisiológica de este proceso es diversa. La bilis se considera principalmente como un secreto digestivo, ya que los ácidos biliares (principalmente sus aniones orgánicos) desempeñan un papel clave en la absorción de grasas. La bilis emulsiona las grasas, aumentando la superficie sobre la cual se hidrolizan bajo la influencia de la lipasa. Bajo la acción de la bilis se encuentra la disolución de los productos de la hidrólisis grasa. Promueve su absorción y resíntesis de triglicéridos en enterocitos. La bilis aumenta la actividad de las enzimas del páncreas y los intestinos (especialmente las lipasas), aumenta la hidrólisis y la absorción de proteínas y carbohidratos.
En violación de la digestión de las grasas, se digieren pobremente y otras sustancias alimenticias, porque la grasa envuelve las partículas más pequeñas de los alimentos y evita la acción de las enzimas sobre ellas. En tales condiciones, la actividad de las bacterias intestinales conduce a un aumento de los procesos de descomposición, fermentación y formación de gas.
Efectos inherentes y reguladores de la bilis: estimulación de la formación de bilis, excreción biliar, actividad motora y secretora del intestino delgado, así como la proliferación y descamación de los enterocitos. La bilis inhibe el proceso de digestión gástrica al neutralizar el ácido que entró en el duodeno e inactivar la pepsina, preparándola para la digestión en el intestino. También tiene un impacto regulatorio en la actividad de evacuación del complejo gastroduodenal. El papel de la bilis en la absorción de vitaminas liposolubles (A, D, E y K), colesterol, sales de calcio de la luz intestinal es importante.
Además, la formación y secreción de bilis se considera una forma peculiar de excreción de ciertas moléculas e iones que no se pueden excretar a través de los riñones. Entre ellos, los más importantes son el colesterol (derivado del colesterol libre, sus ésteres y ácidos biliares), la bilirrubina, así como las moléculas de cobre y hierro. Por lo tanto, la bilis es considerada como un fluido excretor.
La bilis consiste en 80% de agua y 20% de sustancias disueltas en ella. Los últimos incluyen ácidos biliares y sus sales (aproximadamente 65%), fosfolípidos biliares (aproximadamente 20%, principalmente debido a la lecitina), proteínas (aproximadamente 5%), colesterol (4%), bilirrubina conjugada (0,3%), enzimas , inmunoglobulinas, así como numerosas sustancias exógenas y endógenas secretadas con bilis (estirenos vegetales, vitaminas, hormonas, medicamentos, toxinas, iones metálicos, cobre, hierro, potasio, sodio, calcio, zinc, magnesio, mercurio, etc.). En promedio, se segregan 600-700 ml de bilis por día (de 250 a 1500 ml, aproximadamente 10.5 ml por 1 kg de peso corporal). Al mismo tiempo, la secreción de hepatocitos proporciona aproximadamente 500 ml / día del volumen total y alrededor de 150 ml / día se secretan las células de los conductos biliares.
Formacion biliar (coleresis) Continua, y el flujo de bilis hacia el duodeno. (colekinesis) Ocurre periódicamente. Con el estómago vacío, la bilis entra en el intestino de acuerdo con la actividad periódica del hambre. Durante los períodos de descanso, va al WB, donde se concentra, cambia algo su composición y se deposita. Además del agua y las sales, el colesterol y los ácidos grasos libres son absorbidos. En este sentido, distinguen la bilis hepática y vesicular.
La bilis tiene una pequeña actividad enzimática; El pH de la bilis hepática es de 7,3-8,0. A diferencia de los contenidos intestinales, casi no contiene bacterias. Los factores que aseguran la esterilidad de la bilis incluyen la presencia de ácidos biliares (efecto bacteriostático), el rico contenido de inmunoglobulinas, la secreción de moco, la pobreza relativa de los sustratos de energía biliar para las bacterias.
La bilis es una solución micelar. El colesterol, prácticamente insoluble en agua, se transporta en un estado disuelto en la bilis debido a su estructura micelar. Este proceso se denomina disolución coloidal - solubilización.
Los ácidos biliares son moléculas tensioactivas, anfipáticas (hidrofóbicas e hidrófilas) capaces de autoagregarse. En este caso, gracias a una tasa de concentración muy estrecha, llamada concentración crítica de micelización, se forman micelas simples. Las micelas simples tienen una capacidad pronunciada para disolver los lípidos, formando micelas mixtas.
Se cree que las micelas mixtas tienen una estructura cilíndrica: el vástago cilíndrico está lleno de lípidos polares, y las moléculas de ácido biliar están ubicadas entre los extremos polares de las moléculas de lípidos con lados hidrófilos que se enfrentan al entorno del agua, lo que determina su hidrofilicidad (solubilidad en agua).
Las micelas mixtas contienen componentes esenciales: ácidos biliares, localizados en el exterior, fosfolípidos (principalmente fosfatidilcolina, lecitina) y colesterol, ubicados dentro de la micela.
El color de la bilis es de color marrón amarillento debido a la presencia de bilirrubina, la mayor parte de la cual se presenta en forma de bilirrubina diglucuronida, la menor, en forma de bilirrubina monoglucuronida. La bilirrubina asociada no forma parte de las micelas.
Dado que el árbol biliar intercelular es permeable al agua, la vesícula biliar y la bilis hepática son isotónicas.
Formacion biliar
Secreción biliar
Los ácidos biliares son el componente principal de la secreción biliar, se forman solo en el hígado. Ácidos biliares primarios - se sintetizan trihidroxicólicos (colio) y dihidroxicólicos (quenodesoxicólicos) en hepatocitos de colesterol. Ácidos biliares secundarios (desoxicólico y en pequeñas cantidades - litocólico) se forman en el colon a partir del primario como resultado de la modificación bacteriana de los grupos hidroxilo nucleares (7a-deshidroxilación bajo el control de bacterias anaeróbicas). Ácidos biliares terciarios (principalmente ursodeoxicólicos) se forman en el hígado por isomerización de ácidos biliares secundarios.
La síntesis de ácidos biliares a partir del colesterol está regulada por un mecanismo de retroalimentación negativa: un retorno reducido de los ácidos biliares a los hepatocitos con sangre portal conduce a un aumento en su biosíntesis; Cualquier aumento en la síntesis de ácidos biliares se acompaña de un aumento correspondiente en la formación de colesterol.
Los ácidos biliares se conjugan en el hígado con los aminoácidos glicina (alrededor del 80%) o taurina (alrededor del 20%). Cuando se consumen alimentos ricos en carbohidratos, la cantidad de ácidos glicocólicos aumenta en la bilis y en las dietas ricas en proteínas, las taurocólicas. La unión a los aminoácidos evita la absorción de los ácidos biliares en el tracto biliar y en las secciones iniciales del intestino delgado (solo en el íleon terminal y en el intestino grueso). Bajo la acción de las bacterias, es posible la hidrólisis de las sales biliares con la formación de ácidos biliares, glicina o taurina.
Después de la biosíntesis de los ácidos biliares, como se señaló anteriormente, su grupo carboxilo se une al grupo amino de glicina o taurina. Desde un punto de vista químico, tal unión convierte un ácido débil en uno más fuerte. Los ácidos biliares unidos son más solubles a un pH bajo y más resistentes a la precipitación con iones de calcio (Ca 2+) que los ácidos biliares no unidos. Desde el punto de vista biológico, el proceso de unión hace que los ácidos biliares no puedan penetrar a través de las membranas celulares. Por lo tanto, se absorben en los conductos biliares o en el intestino delgado, ya sea en presencia de moléculas portadoras, o si los ácidos biliares experimentan escisión bacteriana. La única excepción a esta regla son los ácidos biliares asociados a glicina dihidroxi, ya que pueden absorberse pasivamente si reciben un ion hidrógeno (H +).
La mayoría de los ácidos biliares (aproximadamente 85-90%), recibidos con el flujo de bilis hacia el intestino delgado, se absorben en la sangre. Sin embargo, la mayoría de los ácidos biliares asociados que se secretan en el intestino delgado se absorben intactos. Una parte más pequeña del ácido no se absorbe intacta, ya que sufre escisión bacteriana en el intestino delgado distal. Se absorben de forma pasiva y, volviendo al hígado, se vuelven a unir y se secretan en la bilis. El 10-15% restante de los ácidos biliares se eliminan del cuerpo principalmente con heces. Esta pérdida de ácidos biliares se compensa con su síntesis en hepatocitos.
Este proceso de escisión intestinal y reabsorción en el hígado es una parte normal del metabolismo de los ácidos biliares. Una pequeña porción de ácidos biliares, no absorbida, entra en el colon. Aquí termina el proceso de división. Además, los ácidos biliares secundarios se forman en el colon bajo la acción de bacterias anaeróbicas (ver arriba).
Los ácidos deoxicólico y litocólico se absorben parcialmente en el colon y vuelven a entrar en el hígado. Después del regreso, el metabolismo de estos ácidos biliares es diferente. El ácido deoxicólico se une a la glicina o taurina y circula con los ácidos biliares primarios. Cabe señalar que en los ancianos, el ácido desoxicólico es el principal ácido biliar en la composición de la bilis. El ácido litocólico no solo se une a la glicina o la taurina, sino que además sulfatos en la posición C-3. Dicha unión "doble" reduce la posibilidad de absorción en el intestino, como resultado de lo cual el ácido litocólico se pierde rápidamente de la composición de los ácidos biliares circulantes; su contenido en la bilis rara vez excede el 5%. La mayoría (95%) de los ácidos biliares, que forman parte de la bilis de los adultos, son ácidos cólicos, quenodesoxicólicos y deoxicólicos.
El proceso de unión de ácidos biliares en hepatocitos es muy efectivo, por lo que los ácidos biliares biliares están completamente presentes en forma conjugada. La posterior escisión bacteriana y la deshidroxilación provocan que los ácidos biliares en las heces se encuentren en estado no unido.
El proceso de deshidroxilación de los ácidos biliares primarios descrito anteriormente reduce su capacidad para disolverse en agua. La formación de ácidos biliares secundarios en los seres humanos prácticamente no tiene importancia fisiológica, a diferencia de los animales. La absorción excesiva de ácido desoxicólico en el colon aumenta el riesgo de cálculos biliares de colesterol. El ácido litocólico se considera hepatotóxico. En experimentos con animales, se demostró que la acumulación de ácido litocólico en el hígado conduce a su derrota. Sin embargo, aún no se ha demostrado que un aumento en la absorción de este ácido en los seres humanos provoque una alteración del hígado.
El ácido ursodesoxicólico, al igual que el ácido desoxicólico, se une al hígado y circula con los ácidos biliares primarios. Sin embargo, la vía metabólica de este ácido es muy corta y el contenido de ácido ursodesoxicólico unido en la bilis nunca excede el 5% de la cantidad total de ácidos biliares. Se cree que la formación de ácido ursodeoxicólico no tiene importancia fisiológica importante.
Los ácidos biliares son poderosos solventes de las grasas, por lo tanto, son citotóxicos en concentraciones que alcanzan la concentración crítica de la micelización. Al mismo tiempo, los ácidos quenodesoxicólicos y desoxicólicos presentes en la bilis son más tóxicos que los asociados con la cólica y el ursodesoxicólico. Aunque los ácidos biliares son demostrativamente citotóxicos in vitro, el epitelio de los conductos biliares y el intestino delgado nunca se daña como resultado de su alta concentración. Esto se asocia, en primer lugar, con la presencia de otros lípidos (fosfolípidos en la bilis y ácidos grasos en el intestino), que reducen la concentración monométrica de los ácidos biliares, y, en segundo lugar, con la presencia de glicolípidos y colesterol en las membranas apicales de las células epiteliales, lo que aumenta la resistencia de las células epiteliales a las dañinas Los efectos de los aniones de ácidos biliares.
Los ácidos cólicos, quenodesoxicólicos y desoxicólicos se absorben y se someten a una circulación enterohepática de 6 a 10 veces por día. El ácido litocólico se absorbe mal y su cantidad en la bilis es pequeña. El conjunto de ácidos biliares es normalmente de aproximadamente 2,5 g, y la producción diaria de ácidos biliares primarios, coloidal y quenodesoxicólico, es en promedio de aproximadamente 330 y 280 mg, respectivamente.
Regulación de la formación de bilis.
Como se sabe, la formación de bilis ocurre continuamente, pero la intensidad de este proceso varía. Mejorar la ingesta de alimentos de formación de bilis y alimentos aceptados El efecto reflejo sobre la coléresis ocurre durante la estimulación de los receptores del tracto gastrointestinal (GIT), órganos internos y efectos reflejos condicionados.
La regulación vegetativa es proporcionada por colinérgicos parasimpáticos (aumentar la formación de bilis) y fibras nerviosas adrenérgicas simpáticas (reducir la formación de bilis).
La regulación humoral se lleva a cabo por la propia bilis debido a la presencia de circulación enterohepática de los ácidos biliares y el mecanismo de retroalimentación negativa (ver arriba). La secretina estimula la secreción de bilis, la secreción de agua y electrolitos en su composición. El glucagón, la gastrina y la colecistoquinina tienen un efecto estimulante más débil.
Excreción biliar
La excreción biliar se considera como una forma peculiar de eliminar las moléculas y los iones del cuerpo que no pueden ser excretados a través de los riñones. Los más importantes entre ellos son el colesterol (como tal y en forma de ácidos biliares) y la bilirrubina, así como los iones de cobre, hierro, etc.
Los principales componentes de la excreción biliar.
El colesterol, prácticamente insoluble en agua, se transporta en la composición de micelas mixtas formadas, como se señaló anteriormente, por los ácidos biliares, los fosfolípidos y el propio colesterol.
Las moléculas de fosfolípidos realizan dos funciones importantes. Primero, aumentan significativamente la solubilidad micelar del colesterol, ya que las micelas mixtas que contienen fosfolípidos solubilizan mucho más colesterol que las micelas simples que contienen solo moléculas de ácido biliar. En segundo lugar, la presencia de fosfolípidos en la bilis reduce la concentración crítica de micelización y la concentración monométrica de ácidos biliares. Como resultado, se reducen la actividad superficial y la citotoxicidad de la bilis hepática.
Con el flujo de bilis, las micelas mixtas entran en el intestino delgado, donde la transformación posterior de sus componentes constitutivos es diferente. Los ácidos biliares disuelven los lípidos, asegurando su absorción, y se absorben en las partes más distales del intestino. Los fosfolípidos biliares, insolubles en agua, se hidrolizan en el intestino y no están involucrados en la circulación enterohepática. Los ácidos biliares regulan su excreción y estimulan la síntesis.
Como se absorbe aproximadamente 1/3 del colesterol, se excreta 2/3 de este. En un adulto, el equilibrio de colesterol está asegurado por su liberación como tal (aproximadamente 600 mg / día) o como ácidos biliares (aproximadamente 400 mg / día). Al mismo tiempo, la circulación enterohepática de los ácidos biliares se considera una forma retardada de excreción de colesterol.
Cabe señalar que en comparación con los animales, el hombre tiene una mayor proporción de colesterol en la bilis. Esto se considera el resultado de la conversión defectuosa del colesterol en ácidos biliares, así como la tasa relativamente baja de secreción de ácidos biliares. Por lo tanto, la bilis está sobresaturada con colesterol en aproximadamente el 25% de las personas de edad avanzada y en el 10-15% de la población se forman cálculos de colesterol (colelitiasis).
Como se señaló anteriormente, la síntesis de ácidos biliares a partir del colesterol está regulada por un mecanismo de retroalimentación negativa.
La bilis es la principal forma de eliminar la bilirrubina del cuerpo. Bilirrubina - El producto final de la descomposición del hemo, en su estructura química es el tetrapirrol. La mayor cantidad de bilirrubina (80-85%) proviene de la hemoglobina en descomposición de los glóbulos rojos que envejecen y de los glóbulos rojos recién formados que colapsan prematuramente en la médula ósea o en el lecho circulatorio (la llamada eritropoyesis ineficaz). El resto de la bilirrubina se forma como resultado de la destrucción de otras proteínas que contienen el dobladillo (por ejemplo, el citocromo P-450, etc.) en el hígado y mucho menos en el tejido extrahepático. Cabe señalar que el catabolismo de la hemoglobina en eritrocitos en la bilirrubina se produce principalmente en los macrófagos del bazo, el hígado y la médula ósea.
Fuentes de formación de bilirrubina (en porcentaje)

Como se sabe, la bilirrubina no conjugada es hidrofóbica (insoluble en agua) y una sustancia potencialmente tóxica que circula en el plasma en un estado estrechamente relacionado con la albúmina y no puede excretarse en la orina. La capacidad del cuerpo para eliminar la bilirrubina se asocia con la eliminación de esta última del plasma sanguíneo por parte de la célula hepática, la posterior conjugación con ácido glucurónico y la liberación en la bilis de un compuesto ya soluble en agua (bilirrubina unida o directa). El proceso de conjugación se realiza a través de la enzima microsomal uridina difosfato glucuronil transferasa. La conjugación de bilirrubina con ácido glucurónico tiene el significado fisiológico más importante, sin embargo, la conjugación de bilirrubina con sulfatos, glucosa y xilosa ocurre en pequeñas cantidades.
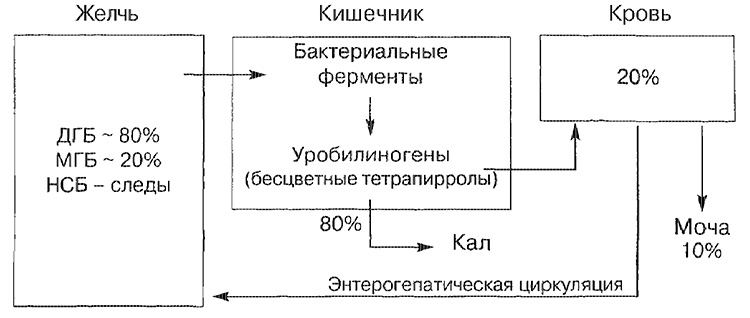
Principales rutas de transporte de bilirrubina.
NSB - bilirrubina no unida; MGB - monoglucurónido de bilirrubina; DGB - bilirrubina diglucuronida.
La secreción de bilirrubina conjugada en los túbulos biliares se produce con la participación de una familia de proteínas de transporte multiespecíficas dependientes de ATP para aniones orgánicos.
La mayoría de la bilirrubina bilirrubina (aproximadamente el 80%) está representada en forma de bilirrubina diglucurónida, una parte más pequeña, en forma de monoglucuronida, y solo una pequeña cantidad está representada por una forma no unida. La bilirrubina, que entró en el intestino, se descompone en la parte terminal del intestino delgado y en el intestino grueso por las enzimas bacterianas (beta-glucuronidasa), que se convierten en tetrapirol incoloro (urobilinógeno). Aproximadamente el 20% de los urobilinógenos formados se reabsorben y necesariamente se excretan en la orina, así como en la bilis (círculo de circulación enterohepático).
La mayoría de los autores están de acuerdo en que la concentración normal de bilirrubina sérica total, determinada por la reacción diazo de Van Der Bergh, por lo general no supera el 1 mg% (0,3-1 mg%, o 5-17 µmol / l). Solo menos del 5% de la bilirrubina se presenta en forma unida. Un aumento en el nivel de bilirrubina en la sangre (hiperbilirrubinemia) y su acumulación en los tejidos conduce a la aparición de ictericia, que, por regla general, se hace notable a valores que superan los 2,5 a 3 mg%.
Como se señaló anteriormente, la bilirrubina circula en la sangre en un estado unido a proteínas y apenas penetra en los fluidos tisulares con un bajo contenido de proteínas. En este sentido, los exudados son de color más amarillo que los transudados. La bilirrubina se adhiere bien al tejido elástico, lo que explica la tinción amarilla temprana de la esclerótica, la piel y las paredes vasculares en la hiperbilirrubinemia. La colestasis prolongada conduce a la aparición de un tono de piel verdoso, que se explica por la deposición de biliverdina.
Será apropiado enfatizar que la presión en los conductos biliares, en un contexto en el que se produce la secreción de bilis, normalmente asciende a 150-200 mm de agua. El art. Su aumento a 350 mm de agua. El art. inhibe la secreción de la bilis, lo que lleva al desarrollo de la ictericia. En el caso de cese completo de la secreción de bilirrubina y ácidos biliares, la bilis se decolora (la llamada bilis blanca).
Además, la bilis es una forma de excreción. grasas vegetales, medicamentos lipófilos y sus metabolitos, diversos xenobióticos que están presentes en las plantas, metabolitos lipófilos, vitaminas liposolubles y hormonas esteroides.
Balanza de hierro y cobre El cuerpo también es compatible debido a la excreción biliar de estos metales. Ambos cationes son secretados en la bilis por una bomba canalicular estimulada por ATP. Con la bilis se produce y la liberación de otros metales.
Excreción de bilis
La corriente de bilis en el sistema biliar es causada por la diferencia de presión en sus diversas secciones y duodeno, tono del esfínter, contracción de las fibras musculares lisas del agujero de gusano y los conductos (las primeras fibras musculares individuales aparecen en los conductos biliares intrahepáticos ubicados en las inmediaciones de las puertas del hígado). La actividad de todo el sistema normalmente está bien coordinada y bajo el control de los mecanismos nerviosos y humorales.
En los cálculos biliares, las concentraciones de bilis se depositan y aumentan con un aumento de 10 veces en la concentración de lípidos, sales biliares, pigmentos biliares, etc. En el momento correcto, se libera en el OZhP y luego en el duodeno. El paso de la bilis coordina al aparato del esfínter. Fuera del proceso digestivo en el duodeno, el esfínter del vial está cerrado. En este momento, el RI y el conducto quístico están relajados, lo que permite que la bilis producida por el hígado ingrese a la luz del RI. La membrana mucosa ZH absorbe agua, iones. Al mismo tiempo la bilis se vuelve más concentrada. Los productos de moco permiten que la bilis esté en estado coloidal.
La presión en el lumen de la glándula en reposo es mucho menor que en los conductos biliares, y es de 60-185 mm de agua. El art. La diferencia en la presión es la base fisiológica del flujo de bilis en la vesícula biliar con el esfínter Oddi cerrado. En el proceso de digestión debido a la reducción de la presión GF aumenta a 150-260 mm de agua. st, asegurando el flujo de la bilis en el duodeno a través de las ampollas del esfínter relajado. Cuando la bilis comienza a fluir hacia el duodeno, la presión en los conductos disminuye gradualmente (con cada comida, el contenido de grasa disminuye 1-2 veces).
El período de la reacción primaria del sistema biliar, debido al tipo, el olor de los alimentos y su recepción, dura entre 7 y 10 minutos. Luego viene el período de evacuación (el principal o el período de vaciado de ZHP), durante el cual, en el contexto de la alternancia de contracciones y relajaciones de la ZP, la vesícula biliar y luego la bilis hepática fluye hacia el duodeno.
La estimulación refleja de la colekinesis (reflejo condicional e incondicional) se produce debido a los receptores ubicados en la cavidad oral, el estómago y el duodeno, a través de los nervios vagos. La regulación humoral se lleva a cabo principalmente bajo la influencia de la colecistoquinina, que tiene un efecto estimulante sobre la FG, lo que provoca su reducción. Además, los cortes débiles en la GI se producen bajo la influencia de la gastrina, la secretina y la Bombezina. Por el contrario, glucagón, calcitonina, anticolincoquinina, péptido vasointestinal y polipéptido pancreático inhiben la reducción de GI.
Las yemas de huevo, las grasas, la leche y la carne son poderosos estimulantes de la excreción de bilis.
Los trastornos del aparato del esfínter y la motilidad gonadal pueden causar cambios en la dirección y la velocidad del flujo biliar, la disquinesia, el reflujo de las secreciones pancreáticas y el contenido de duodeno en la ACP, y la bilis en el conducto pancreático, que es la base para el desarrollo de una serie de estados patológicos y enfermedades.
Las sales biliares, la bilirrubina conjugada, el colesterol, los fosfolípidos, las proteínas, los electrolitos y el agua son secretados por los hepatocitos hacia los canalículos biliares. Aparato de secreción de bilis incluye proteínas de transporte. membrana tubular, orgánulos intracelularesy estructuras citoesqueleto. Contactos estrechosentre los hepatocitos, el lumen de los túbulos se separa del sistema circulatorio del hígado.
La membrana tubular contiene proteínas de transporte para ácidos biliares, bilirrubina, cationes y aniones. Microvilli aumenta su área. Los orgánulos están representados por el aparato de Golgi y los lisosomas. Con la ayuda de vesículas, las proteínas (p. Ej., IgA) se transportan desde la membrana sinusoidal a la membrana canalicular, el transporte de las proteínas de transporte sintetizadas en la célula para el colesterol, los fosfolípidos y, posiblemente, los ácidos biliares de los microsomas a la membrana tubular.
El citoplasma del hepatocito alrededor del túbulo contiene la estructura del citoesqueleto: microtúbulos, microfilamentosy filamentos intermedios.
Los microtúbulos se forman por polimerización de tubulina y forman una red dentro de la célula, especialmente cerca de la membrana basolateral y el aparato de Golgi, que participan en el transporte vesicular mediado por receptores, la secreción de lípidos y, en ciertas condiciones, los ácidos biliares. La formación de microtúbulos es inhibida por la colchicina.
Las actinas interactivas polimerizadas (F) y libres (G) participan en la construcción de microfilamentos. Los microfilamentos, concentrados alrededor de la membrana tubular, determinan la contractilidad y la motilidad de los túbulos. La faloidina, que aumenta la polimerización de la actina, y la citocalasina B, que la debilita, inhiben la motilidad de los túbulos y causan colestasis.
Los filamentos intermedios están formados por citoqueratinas y forman una red entre las membranas plasmáticas, el núcleo, los orgánulos intracelulares y otras estructuras del citoesqueleto. La ruptura de los filamentos intermedios conduce a la interrupción de los procesos de transporte intracelular y la obliteración de la luz de los túbulos.
El agua y los electrolitos afectan la composición de la secreción tubular, penetrando a través de contactos estrechos entre los hepatocitos debido al gradiente osmótico entre la luz tubular y los espacios de Disse. (corriente paracelular).La integridad de los contactos estrechos depende de la presencia en la superficie interna de la membrana plasmática de la proteína ZO-1 con una masa molecular de 225 kDa. La ruptura de los contactos estrechos se acompaña de la entrada de moléculas más grandes disueltas en los canalículos, lo que conduce a la pérdida del gradiente osmótico y al desarrollo de colestasis. Al mismo tiempo, se puede observar regurgitación de la bilis canalicular en la sinusoide.
Los canalículos biliares fluyen hacia los conductos, a veces llamados colangioles o canales de Goering. Las ductulas se localizan principalmente en las áreas portales y fluyen hacia los conductos biliares interlobulares, que son el primero de los tractos biliares acompañados por ramas de la arteria hepática y la vena porta, y se encuentran en la composición de las tríadas portales. Los conductos interlobulares, que se fusionan, forman conductos septales hasta que emergen dos conductos hepáticos principales, que emergen de los lóbulos derecho e izquierdo en la fisura portal del hígado.
Secreción de bilis
La formación de bilis ocurre con la participación de una serie de procesos de transporte volátiles. Su secreción es relativamente independiente de la presión de perfusión. La corriente total de bilis en humanos es de aproximadamente 600 ml / día. Los hepatocitos proporcionan la secreción de dos fracciones de bilis: dependen de los ácidos biliares ("225 ml / día) y no dependen de ellos (" 225 ml / día). Los 150 ml / día restantes son secretados por las células del conducto biliar.
La secreción de sales de ácidos biliares es el factor más importante en la formación de bilis. (fracción dependiente de los ácidos biliares).El agua se mueve a raíz de las sales osmóticamente activas de los ácidos biliares. Los cambios en la actividad osmótica pueden regular el flujo de agua hacia la bilis. Existe una clara correlación entre la secreción de sales de ácidos biliares y el flujo de bilis.
La existencia de la fracción biliar, que no depende de los ácidos biliares, se demuestra por la posibilidad de la formación de bilis que no contiene sales de ácidos biliares. Por lo tanto, es posible continuar el flujo de bilis, a pesar de la ausencia de excreción de sales de ácidos biliares; La secreción de agua en este caso se debe a otras sustancias solubles osmóticamente activas, como el glutatión y los bicarbonatos.
Mecanismos celulares de la secreción biliar.
Un hepatocito es una célula epitelial secretora polar que tiene membranas basolaterales (sinusoidales y laterales) y apicales (tubulares).
La formación de bilis incluye la captura de ácidos biliares y otros iones orgánicos e inorgánicos, su transporte a través de la membrana basolateral (sinusoidal), citoplasma y membrana tubular. Este proceso se acompaña de la filtración osmótica del agua contenida en el espacio de los hepatocitos y paracelulares. La identificación y caracterización de las proteínas de transporte de las membranas sinusoidales y tubulares es compleja. Especialmente difícil es el estudio del aparato secretor de los túbulos; sin embargo, hasta la fecha, una técnica para obtener hepatocitos duplicados en un cultivo de corta duración se ha desarrollado y demostrado ser confiable en muchos estudios. La clonación de proteínas de transporte nos permite caracterizar la función de cada uno de ellos por separado.
El proceso de formación de bilis depende de "la presencia de ciertas proteínas portadoras en las membranas basolaterales y tubulares. Na +, K desempeña el papel de la fuerza impulsora de la secreción + - ATPasa de la membrana basolateral, que proporciona un gradiente químico y una diferencia de potencial entre el hepatocito y el espacio circundante. Na +, K + - La ATPasa intercambia tres iones de sodio intracelular por dos iones de potasio extracelulares, manteniendo un gradiente de concentración de sodio (alto exterior, bajo interior) y potasio (bajo exterior, alto interior). Como resultado, el contenido celular tiene una carga negativa (–35 mV) en comparación con el espacio extracelular, lo que facilita la captura de iones cargados positivamente y la excreción de iones cargados negativamente. Na +, K + -ATPase no se detecta en la membrana tubular. El flujo de membrana puede afectar la actividad enzimática.
Captura en la superficie de una membrana sinusoidal.
La membrana basolateral (sinusoidal) tiene muchos sistemas de transporte para capturar aniones orgánicos, cuya especificidad de sustrato se superpone parcialmente. La caracterización de las proteínas portadoras se dio previamente sobre la base del estudio de las células animales. La reciente clonación de proteínas de transporte humano ha permitido caracterizar mejor su función. La proteína de transporte para los aniones orgánicos (proteína transportadora de aniones orgánicos - OATP) es independiente del sodio, contiene moléculas de varios compuestos, incluidos ácidos biliares, bromsulfaleína y, probablemente, bilirrubina. Se cree que el transporte de bilirrubina en el hepatocito también es realizado por otros portadores. La captura de ácidos biliares conjugados con taurina (o glicina) se realiza mediante la proteína transportadora de sodio / taurocolato (proteína cotransportadora de sodio / ácido biliar - NTCP).
La transferencia de iones a través de la membrana basolateral implica una proteína que intercambia Na + / H + y regula el pH dentro de la célula. Esta función también es realizada por la proteína cotransport para Na + / HCO 3 -. En la superficie de la membrana basolateral, también se capturan sulfatos, ácidos grasos no esterificados y cationes orgánicos.
Transporte intracelular
El transporte de ácidos biliares en el hepatocito se lleva a cabo utilizando proteínas citosólicas, entre las cuales la Z-hidroxiesteroide deshidrogenasa desempeña el papel principal. La glutatión-S-transferasa y las proteínas de unión a ácidos grasos son de menor importancia. El retículo endoplásmico y el aparato de Golgi están involucrados en la transferencia de ácidos biliares. Aparentemente, el transporte vesicular se activa solo cuando los ácidos biliares se inyectan significativamente en la célula (en concentraciones que exceden la fisiológica).
El transporte de proteínas en la fase líquida y los ligandos, como la IgA y las lipoproteínas de baja densidad, se realiza mediante la transcitosis vesicular. El tiempo de transferencia desde la membrana basolateral a la tubular es de aproximadamente 10 minutos. Este mecanismo es responsable solo por una pequeña parte de la corriente total de la bilis y depende del estado de los microtúbulos.
Secreción tubular
La membrana tubular es una sección especializada de la membrana plasmática de hepatocitos que contiene proteínas de transporte (en su mayoría dependientes de ATP) responsables de la transferencia de moléculas a la bilis contra un gradiente de concentración. Las enzimas como la fosfatasa alcalina, GGTP también se localizan en la membrana canalicular. La transferencia de glucurónidos y glutatión-S-conjugados (por ejemplo, bilirrubina diglucurónido) se realiza con la ayuda de una proteína de transporte multispecífico canalicular para aniones orgánicos (glándula cMOAT); transporter - SWAT), cuya función está parcialmente controlada por un potencial intracelular negativo. La corriente de bilis, que no depende de los ácidos biliares, está aparentemente determinada por el transporte de glutaión, así como por la secreción canalicular de bicarbonato, posiblemente con la participación de una proteína que intercambia Cl - / HCO 3 -.
Un papel importante en el transporte de sustancias a través de la membrana tubular pertenece a dos enzimas de la familia de las glicoproteínas P; ambas enzimas son dependientes de ATP. Proteína de resistencia a múltiples fármacos 1 (MDR1) La proteína de resistencia a múltiples fármacos 1 transfiere cationes orgánicos y también elimina los fármacos citostáticos de las células cancerosas, causando su resistencia a la quimioterapia (de ahí el nombre de proteína). El sustrato endógeno de MDR1 es desconocido. MDR3 transporta fosfolípidos y actúa como una flipasa para la fosfatidilcolina. La función de MDR3 y su importancia para la secreción de fosfolípidos en la bilis se refinan en experimentos en ratones que carecen de la glucoproteína mdr2-P (análoga a la MDR3 humana). En ausencia de fosfolípidos en la bilis, los ácidos biliares causan daños en el epitelio biliar, inflamación de los conductos y fibrosis periductular.
El agua y los iones inorgánicos (especialmente el sodio) se excretan en los capilares biliares a lo largo del gradiente osmótico por difusión a través de contactos apretados semipermeables cargados negativamente.
La secreción de bilis está regulada por muchas hormonas y mensajeros secundarios, incluidos el AMPc y la proteína quinasa C. El aumento de la concentración de calcio intracelular inhibe la secreción de bilis. El paso de la bilis a través de los túbulos se debe a los microfilamentos que proporcionan motilidad y contracción de los túbulos.
Secreción dúctil
Las células epiteliales de los conductos distales producen un secreto rico en bicarbonato que modifica la composición de la bilis tubular (la llamada corriente ductular, bilis).El proceso de secreción produce AMPc, algunas proteínas de transporte de membrana, incluida una proteína que intercambia Cl - / HCO 3 - y regulador de la conducción transmembrana en la fibrosis quística -membrana canal para Cl -, cAMP ajustable. La secreción ductular es estimulada por la secretina.
Se asume que el ácido ursodesoxicólico es absorbido activamente por las células ductulares, intercambiado por bicarbonatos, reciclado en el hígado y posteriormente excretado nuevamente en la bilis ("derivación colohepática"). Quizás esto explica el efecto colerético del ácido ursodeoxicólico, acompañado de una alta secreción de bicarbonato biliar en la cirrosis experimental.
La presión en los conductos biliares, en la que se produce la secreción de bilis, es normalmente de 15 a 25 cm de agua. El art. Aumentar la presión a 35 cm de agua. El art. Conduce a la supresión de la secreción biliar, desarrollo de ictericia. La secreción de bilirrubina y ácidos biliares se puede detener completamente, mientras que la bilis se vuelve incolora (hiel blanca)y se asemeja al fluido mucoso.
La bilis es un líquido de plasma isosmótico que consiste en agua, electrolitos y sustancias orgánicas (ácidos biliares, fosfolípidos, colesterol, bilirrubina). Los ácidos biliares (o sus sales) son el principal componente orgánico de la bilis. Los ácidos biliares entran en la bilis de dos fuentes: (1) ácidos biliares primarios (cólicos y quenodesoxicólicos), que se sintetizan a partir del colesterol en el hígado; (2) Los ácidos biliares secundarios (desoxicólicos, litocólicos y ursodesoxicólicos) se forman por la acción de bacterias intestinales de los ácidos biliares primarios. Los propios ácidos biliares constan de dos componentes importantes que determinan sus propiedades fisiológicas y fisicoquímicas: (1) un núcleo de esteroides con sustituyentes hidroxilo; (2) una cadena lateral alifática (Figura 7-4).
La figura 7-4.
Los ácidos biliares constan de dos componentes: un núcleo con extremos hidroxilo y una cadena lateral alifática. En la figura, el ácido cólico se muestra como un ejemplo de ácido 3-hidroxi (3 \\ beta -, 7 \\ beta -, 12 \\ alpha - OH). Otros ejemplos son los ácidos biliares que contienen desoxicolato (3 \\ beta -, 12 \\ beta - OH), quenodeoxicolato (3 \\ beta -, 7 \\ beta - OH) y litocolato (3 \\ alpha - OH).
En la mayoría de los mamíferos, los ácidos biliares primarios contienen de tres a siete sustituyentes hidroxilo, cuyo número afecta a su solubilidad en agua (hidrofilicidad). Poco después de la formación, los ácidos biliares primarios se modifican en el grupo carboxilo terminal. Esto ocurre durante la fase hepática de la circulación enterohepática de los ácidos biliares secundarios y su conjugación con glicina o taurina. La presencia de componentes hidrófilos (componentes hidroxilo y enlaces amida de la cadena lateral alifática) e hidrófobos (núcleo esteroide) permite que las moléculas de ácidos biliares conjugados actúen como un compuesto anfótero. Esto les da la oportunidad de formar micelas (agregados polimoleculares) por encima de la concentración micelar crítica. A su vez, las moléculas de los ácidos biliares son capaces de disolver otras sustancias anfóteras (colesterol, fosfolípidos) con la formación de micelas mixtas. Este papel similar al detergente de los ácidos biliares es importante para la estabilización del estado físico-químico de la bilis, la digestión y la absorción de las grasas.
La síntesis de ácidos biliares a partir del colesterol está regulada por un mecanismo de retroalimentación negativa, aunque la naturaleza de la regulación a nivel molecular y bioquímico aún no se conoce completamente. La 7? -Hidroxilación microsomal del colesterol es un paso crítico en la síntesis de ácidos biliares. El ácido chenodesoxicólico, utilizado para disolver los cálculos de la vesícula biliar, inhibe la síntesis de ácidos biliares y, por lo tanto, aumenta los niveles de colesterol en la sangre. Cuando se usa ácido ursodeoxicólico, tales cambios no se observan incluso con el tratamiento a largo plazo.
La formación de bilis tiene lugar tanto en la superficie sinusoidal como en la tubular de la membrana del hepatocito y es un proceso tanto intracelular como paracelular. En contraste con la filtración glomerular en los riñones, que pasivamente bajo la acción de las fuerzas hidrostáticas, durante la formación de la bilis, hay una transferencia activa de componentes orgánicos e inorgánicos hacia el lumen de los túbulos y el transporte pasivo de agua. Por lo tanto, los procesos de secreción de bilis son similares a los procesos de secreción en el acini del páncreas, el epitelio de los túbulos renales. La formación de bilis tubular se puede dividir en dos tipos (fig. 7-5): (1) formación de bilis, según la secreción de ácidos biliares, definida como la proporción de la cantidad de bilis secretada en los túbulos a la cantidad de sales biliares secretadas; (2) formación de bilis, independiente de la secreción de ácidos biliares, que se puede representar como una secreción activa de electrolitos inorgánicos y otras sustancias y se refleja en el gráfico como una intersección en y de esta línea. En otras palabras, la formación de bilis asociada con la secreción ácida es el flujo de la bilis, dependiendo de la presencia de sales biliares osmóticamente activas en los conductos biliares, y la formación de bilis no relacionada con la secreción ácida en ausencia de sales biliares. La proporción de la tasa de formación de bilis y la formación de sales de ácidos biliares no es lineal con pequeñas cantidades de bilis secretada y no puede corresponder a la relación lineal mostrada en la Fig. 7-5. Por lo tanto, ambos tipos de formación de bilis deben considerarse como indicadores interrelacionados de formación de bilis.
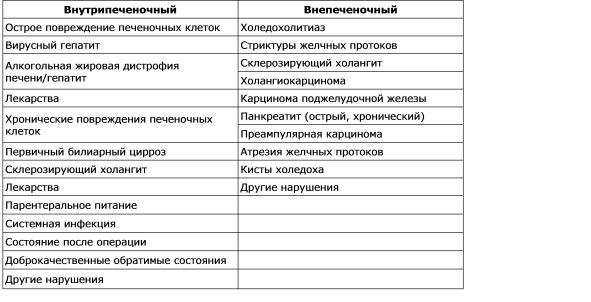
La violación de la formación de la bilis se llama colestasis. La secuencia de las manifestaciones patológicas, fisiológicas y clínicas emergentes de la colestasis depende de su causa. Se detecta un estudio morfológico del material de la bilis hepatobiopsia en los túbulos de los hepatocitos pericentral, se observa la dilatación de los túbulos y el estudio de la ultraestructura revela una disminución en el número de microvilos. La colestasis se puede definir como un defecto funcional en la formación de la bilis a nivel del hepatocito (colestasis intrahepática), así como la secreción y salida orgánica o mecánica deteriorada de la bilis (colestasis extrahepática). Las causas más comunes de colestasis intrahepática y extrahepática se dan en la Tabla. 7-2. Hay varios mecanismos que juegan un papel importante en la patogénesis de la colestasis intrahepática: deterioro y daño a la función de la membrana sinusoidal; alteración de las funciones de los orgánulos intracelulares de hepatocitos; Daño y rotura de la membrana tubular. Por lo tanto, no existe un mecanismo único para la colestasis en diversas situaciones clínicas, y la multiplicidad de mecanismos puede conducir a diversos trastornos. Clínicamente, la colestasis se caracteriza por un aumento en los niveles sanguíneos de muchas sustancias, como la bilirrubina, las sales biliares y el colesterol, que normalmente se secretan en la bilis. En el análisis bioquímico de la sangre con colestasis, se observa un aumento desproporcionado en la actividad de la fosfatasa alcalina y la concentración de bilirrubina en paralelo con los cambios en el nivel de aminotransferasas, que se explicarán más adelante.
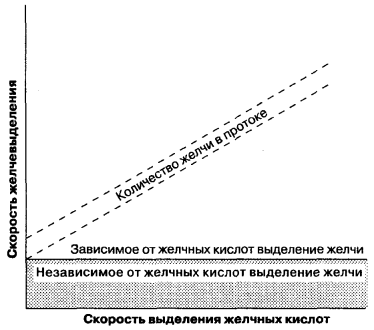
La figura 7-5.
Formación de bilis asociada con la secreción de ácidos biliares, formación de bilis independiente de la secreción de ácidos biliares. (Por: Moseley R. H., Secreción de bilis. En: Yamada T., Alpers D. H., Owyang C., Powell D. W., Silverstein F. E., editores. Libro de texto de Gastroenterología, 2ª edición. Filadelfia: J. B. Lippincott, 1995: 387.)
Tabla 7 - 2.
En el hígado, se forma el jugo digestivo más importante - la bilis.
La bilis es producida por los hepatocitos mediante el transporte activo y pasivo de agua, colesterol, bilirrubina y cationes en ellos. En los hepatocitos a partir del colesterol, se forman ácidos biliares primarios, cholic y deoxycholic. Un complejo soluble en agua se sintetiza a partir de bilirrubina y ácido glucurónico. Entran en los capilares y conductos biliares, donde los ácidos biliares se combinan con la glicina y la taurina. Como resultado, se forman ácidos glicocólicos y taurocólicos. El bicarbonato de sodio se forma por los mismos mecanismos que en el páncreas.
La bilis es producida por el hígado todo el tiempo. En su día se forma alrededor de 1 litro. Los hepatocitos excretan la bilis primaria o hepática. Este líquido es una reacción alcalina de color amarillo dorado. Su pH = 7.4 - 8.6. Se compone de 97.5% de agua y 2.5% de sólidos. El residuo seco contiene:
1. Sustancias minerales. Sodio, potasio, cationes de calcio, bicarbonato, fosfato, aniones de cloro.
2. Ácidos ácidos - taurocólicos y glicocólicos.
3. Pigmentos biliares - bilirrubina y su forma oxidada biliverdina. La bilirrubina da color a la bilis.
4. Colesterol y ácidos grasos.
5. Urea, ácido úrico, creatinina.
Dado que fuera del sistema digestivo, el esfínter de Oddi, ubicado en la boca del conducto biliar común, está cerrado, la bilis excretada se acumula en vesícula biliar. Aquí el agua se reabsorbe, y el contenido de componentes orgánicos básicos y mucina aumenta de 5 a 10 veces. Por lo tanto, la bilis quística contiene 92% de agua y 8% de residuo seco. Es más oscuro, más grueso y más viscoso que el hígado. Debido a esta concentración, la vejiga puede acumular bilis durante 12 horas. Durante la digestión, el esfínter de Oddi y el esfínter de Lutkens en el cuello de la vejiga se abren. La bilis entra en el duodeno.
Valor de la bilis:
1. Los ácidos málicos emulsionan una parte de las grasas, convirtiendo partículas grandes de grasa en gotas finas.
2. Activa las enzimas del jugo intestinal y pancreático, especialmente la lipasa.
3. En combinación con los ácidos biliares, la absorción de los ácidos grasos de cadena larga y las vitaminas liposolubles se produce a través de la membrana de los enterocitos.
4. Zhelchka promueve la resíntesis de triglicéridos en enterocitos.
5. Inactiva las pepsinas, y también neutraliza el quimo agrio que proviene del estómago. Esto asegura la transición de la digestión gástrica a la intestinal.
6. Estimula la secreción de jugos pancreáticos e intestinales, así como la proliferación y descamación de enterocitos.
7. Fortalece la motilidad intestinal.
8. Tiene un efecto bacteriostático en los microorganismos intestinales y, por lo tanto, evita el desarrollo de procesos putrefactos en él.
La regulación de la formación de bilis y la excreción biliar se realiza principalmente por mecanismos humorales, aunque los nerviosos desempeñan un cierto papel. El estimulador más poderoso de la formación de bilis en el hígado son los ácidos biliares, absorbidos en la sangre desde el intestino. También se ve reforzada por la secretina, que contribuye a un aumento en el bicarbonato de sodio en la bilis. El nervio vago estimula la producción de bilis, la inhibición simpática.
Cuando el quimo entra en el duodeno, las células I comienzan a liberar sus células i de colecistoquinina-pancreozymin. Especialmente este proceso es estimulado por grasas, yema de huevo y sulfato de magnesio. CCK-PZ fortalece las contracciones de los músculos lisos de la vejiga, los conductos biliares, pero relaja los esfínteres de Lutkens y Oddi. La bilis se libera en el intestino. Los mecanismos reflejos juegan un pequeño papel. El quimo irrita los quimiorreceptores del intestino delgado. Los impulsos de ellos entran en el centro digestivo de la médula oblonga. De él están en el vago al tracto biliar. Los esfínteres se relajan y los músculos lisos de la vejiga se contraen. Promueve la excreción biliar.
Las enfermedades más graves son la hepatitis y la cirrosis. Más a menudo, la hepatitis es el resultado de una infección (hepatitis infecciosa A, B, C) y la exposición a productos tóxicos (alcohol). En la hepatitis, los hepatocitos se ven afectados y todas las funciones del hígado están afectadas. La cirrosis es el resultado de la hepatitis. La violación más común de la excreción biliar es la colelitiasis. La mayor parte de los cálculos biliares están formados por el colesterol, ya que la bilis de estos pacientes está supersaturada con ellos.
El hígado es el órgano más grande del cuerpo y es fundamental para el metabolismo. Desempeña muchas funciones, participando en el metabolismo de proteínas, carbohidratos, grasas, hormonas y vitaminas, así como en la neutralización de muchas sustancias endógenas y exógenas. Estos procesos se describen en los libros de texto de química fisiológica y en esta sección en relación con sistema digestivo consideraremos solamente función hepática excretoraes decir, secreción de bilis. Bilis consiste en agua, sales minerales, moco, lípidos de colesterol y lecitina y dos tipos de componentes específicos: ácidos biliares y pigmento de bilirrubina. Los ácidos biliares son detergentes y su acción emulsionante juega un papel importante en la digestión de los lípidos. La bilirrubina es el producto final de la descomposición de la hemoglobina que se eliminará del cuerpo.
^
Formacion biliar
Anatomía funcional.Células hepáticas (hepatocitos)formar una placa en el espesor de una celda, separada por rendijas estrechas (Espacio de disse)representando lo completado
^ 766 PARTE VIII. ALIMENTACIÓN, DIGESTIÓN Y EXTRACTO.
Por sangre sinusoidesequivalente a los capilares. En las paredes de los sinusoides hay poros a través de los cuales pueden pasar macromoléculas tan grandes como la albúmina y las lipoproteínas. En los mas pequeños túbuloscapilares biliares limitados por las membranas plasmáticas de dos hepatocitos adyacentes: la bilis se recoge en grandes canalículos de Goring,las paredes de las cuales, como las más grandes. túbulo intercolegioy conductos biliaresformado por células secretoras cúbicas. Los pequeños túbulos dentro de los lóbulos del hígado y entre ellos se funden en otros más grandes, formando finalmente conducto hepático.De este conducto sale. conducto císticoa la vesícula biliar. Tras la fusión, se forman los conductos hepáticos y quísticos. conducto biliar comúnapertura en el duodeno en la parte superior de la papila del Vater, detrás o cerca del conducto pancreático (Fig. 29.1).
^ Funciones de la bilis.La bilis realiza muchas funciones importantes. Con ella los productos finales se muestranintercambio, como la bilirrubina, así como las drogas y toxinas. Asignación conbilis colesteroljuega un papel importante en la regulación de su equilibrio. Ácidos biliaresesencial para la emulsificación y absorción de grasas. Además, la bilis contiene agua, sales minerales y mucosidad. Cerca de 24 horas al día. 600 mlbilis, y 2/3 de esta cantidad proviene de los túbulos y 1/3 de los conductos más grandes.
^ Bilis tubular se forma en cantidades aproximadamente iguales con la participación de dos mecanismos diferentes, dependientes de la bilis y dependientes de la bilis (fig. 29.27).
^ Secreción dependiente de ácidos biliares. Hay una estrecha relación entre tasa de flujo biliary secreción de ácidos biliares.En la bilis canalicular, la concentración de ácidos biliares es 100 veces mayor que en la sangre portal, por lo tanto, se considera que son secretados por transporte activo que involucra vectores.Después de los ácidos biliares a lo largo del gradiente osmótico, el agua se precipita en los túbulos, por lo que la bilis es isotónica con la sangre.
Hay dos fuentes de ácidos biliares. Primero, se sintetizan de novo a partir del colesterol en los propios hepatocitos con la participación de 7-hidroxilasa. Esta enzima desempeña un papel clave y controla la velocidad de síntesis de los ácidos biliares a través de un mecanismo de retroalimentación. En segundo lugar, los hepatocitos son capaces de absorber activamente los ácidos biliares de sangre portaly excretarlos en túbulos (ver también fig. 29.29). Esta extracción es muy efectiva; Con un solo paso de sangre a través del hígado, el 80% de los ácidos biliares se extraen de él. Por lo tanto
La concentración de ácidos biliares en la sangre periférica es mucho menor que en el sistema portal. Dado que los ácidos biliares se extraen de la sangre 6 veces más rápido de lo que ingresan a los túbulos, este último proceso es el que limita la tasa de secreción de ácidos biliares.
^ Secreción independiente de ácidos biliares. En
Este proceso involucra iones Na +, Cl -, HCO 3 - y agua. La fuerza motriz es transporte activo Να + , posiblemente con bicarbonato. Una secreción independiente de ácidos biliares estimula, en particular, secretin
Además de los ácidos biliares, la bilirrubina, el colesterol y los fosfolípidos (principalmente lecitina) se secretan activamente en los canalículos (Fig. 29.27). Insoluble en agua ("Indirecto") bilirrubina,la mayoría de los cuales se forma a partir de la hemoglobina de los glóbulos rojos de edad, ingresa a los hepatocitos en forma de un agregado coloidal asociado con la albúmina. Su formación diaria es de aproximadamente 4 g / kg de peso corporal, o 200-300 mg / día. En los hepatocitos, el 80% de la bilirrubina está conjugada. conÁcido glucurónico y una pequeña cantidad con ácido sulfúrico. En tales conjugados
^ CAPÍTULO 29. FUNCIONES DEL TRACTO GASTROINTESTINAL 767
La bilirrubina se excreta en la bilis. (Bilirrubina "directa").En general, las drogas y toxinas se eliminan de la misma manera.
^ Modificación de la bilis en los conductos biliares.
(fig. 29.27). En los conductos donde se abren los túbulos, se modifica la bilis primaria. Este proceso se asemeja al proceso de modificar el filtrado glomerular en los túbulos renales (pág. 785), y de manera similar se calcula liquidaciónsolo en el caso de la bilis como sustancia inerte se usa en lugar de inulina. eritritolo manitolque se secretan en los túbulos, pero no se reabsorben. Estudios similares han demostrado que aproximadamente 180 ml de bilis, o 1/3 de su cantidad total, se secretan en los conductos con la secreción activa de NSO ^ ". Este proceso se estimula secretin
^
Bilis hepática y vesicular.
La composición de la bilis hepática.(tab. 29.3). La bilis secretada por el hígado a una velocidad de 0,4 ml / min tiene un color dorado, que se explica por la presencia de bilirrubina. La concentración de electrolitos en esta bilis es la misma que en el plasma, excepto por el hecho de que contiene el doble de HCO3 "y algo menos que el CI ~. Al mismo tiempo, la composición de las bilis de sustancias orgánicas es muy diferente del plasma, ya que en la bilis son Representado casi exclusivamente por ácidos biliares, colesterol y fosfolípidos.
 |
^ Ácidos biliaresse forman en el hígado a partir del colesterol como resultado de su hidroxilación y la adición de un grupo carboxilo. Los ácidos que se forman en el hígado son ácidos biliares primarios;estos incluyen chenodeoxycholic(dioxiforme) y holevaya(trioxyforma) acidoEn el hígado, no están en forma libre, sino en forma de conjugados con glicina y taurina, y los conjugados con glicina se forman tres veces más, ya que la cantidad de taurina es limitada. Los ácidos biliares conjugados son más solubles en agua,
Que no están conjugados, y tienen una mayor capacidad para disociarse y formar sales biliares con cationes, principalmente con iones Na +. En un ambiente ácido (pH 4.0), las sales de ácidos biliares son insolubles y precipitadas, pero a valores de pH fisiológicos (en el intestino delgado), son muy solubles.
En la parte distal del íleon y en el colon, algunas de las sales de los ácidos biliares primarios se someten a deshidroxilación bajo la acción de bacterias anaeróbicas y se convierten en ácido biliar-litocólico secundario(monooxyform) y desoxicólico(dioxiforme). Los ácidos chenodeoxicólicos, cólicos y desoxicólicos están presentes en una proporción de 2: 2: 1. El ácido litocólico está presente solo en algunas fracciones, ya que la mayor parte se excreta.
El efecto emulsionante de los ácidos biliares en las grasas se basa principalmente en su capacidad para formar micelas.Las moléculas de ácido biliar tienen una estructura tridimensional en la que los grupos hidrófilo carboxilo e hidroxilo están en el mismo lado de la molécula, y la parte hidrófoba de la molécula (núcleo esteroide, grupos metilo) está en el lado opuesto, debido a que las moléculas de ácido biliar tienen y hidrofilicoy propiedades lipofílicas.Debido a esta estructura, las moléculas de los ácidos biliares actúan como detergentes: en la interfaz de las fases lipídica y acuosa, forman una película casi monomolecular en la que los grupos hidrófilos se convierten en la fase acuosa y la fase lipófila en la lipídica. En la fase acuosa, los ácidos biliares forman agregados ordenados. -micelossiempre que su concentración alcance un cierto nivel, llamado concentración micelar crítica(1-2 mmol / l). La región lipófila interna de la micela puede contener lípidospor ejemplo, colesterol y fosfolípidos; Estas micelas se llaman mixtas (Fig. 29.28). El colesterol en sí mismo es insoluble en agua, pero puede estar en solución en la composición de las micelas. Si su concentración excede la capacidad de las micelas, forma un precipitado cristalino; este proceso subyace a la formación de cálculos biliares de colesterol (pág. 769).
^ La composición de la bilis quística. (tab. 29.3). La capacidad de la vesícula biliar es de solo 50-60 ml. Al mismo tiempo, el hígado secreta bilis a una velocidad de 600 ml / día, y la mitad de esta cantidad pasa a través de la vesícula biliar antes de entrar en el intestino delgado. Se compensa la diferencia entre el volumen de bilis que ingresa a la vesícula biliar y su capacidad. reabsorción altamente eficienteagua en la vesícula biliar. En unas pocas horas, el 90% del agua puede ser reabsorbida por la bilis. Con
^
768 PARTE VIII. ALIMENTACIÓN, DIGESTIÓN Y EXTRACTO.
Esta materia orgánica permanece en la vesícula biliar y aumenta su concentración en la bilis. La fuerza motriz de la reabsorción es transporte activo de iones Nα *con la participación de una "bomba" incrustada en las membranas basales y laterales de las células y activada por (Na + -K +) - ATPasa. Siguiendo los iones Na +, los iones Cl - y HCO 3 - se difunden en la dirección del gradiente eléctrico o se transportan por los transportistas. Como resultado de la reabsorción de HCO3, el pH de la vesícula biliar disminuye a 6.5 frente a 8.2 en la bilis hepática. Como resultado de la creación de una alta concentración de iones Na + en el epitelio de la vesícula biliar en el espacio extracelular, se produce un gradiente osmótico que conduce al bombeo de agua, que luego fluye hacia los capilares (pág. 751).
^ Motilidad de la vesícula biliar. En el estado de ayuno, la bilis se acumula en la vesícula biliar, y en
El tiempo de comer se asigna como resultado de las contracciones de la vesícula biliar. El principal estimulador de la actividad contráctil de la vesícula biliar es. colecistoquinina,segregado por la membrana mucosa del duodeno al ingresar a él de grasa que contiene quimo. Las contracciones de la vesícula biliar también se estimulan en cierta medida. nervio errantey parasimpatolíticos.Comienzan a los 2 minutos después de que los alimentos grasos entren en contacto con la mucosa intestinal, y después de 15 a 90 minutos, la vejiga se vacía por completo. La motilidad de la vesícula biliar implica dos procesos. Inicialmente, se desarrolla una contracción tónica, como resultado de lo cual el diámetro de la vesícula biliar disminuye, y luego se imponen contracciones periódicas sobre este efecto, cuya frecuencia es de 2-6 / min. Como resultado de estos dos procesos, se crea una presión de 25–30 mm Hg. El art.
^
Circulación enterohepática
Circulación de ácidos biliares(fig. 29.29). Los ácidos biliares se secretan en el duodeno como micelas mixtas. A pesar de la dilución de los ácidos biliares por el contenido del estómago, su concentración en el intestino es de aproximadamente 10 mmol / ly permanece por encima de la concentración crítica de formación de micelas. Aquí, además del colesterol y la lecitina, las micelas incluyen los productos de la descomposición de las grasas hidrolíticas: acidos grasosy monogliceridosEn el contacto inicial de las micelas con la pared intestinal.
^
CAPÍTULO 29. FUNCIONES DEL TRACTO GASTROINTESTINAL 769
Los lípidos se difunden a través de la membrana del borde en cepillo a los enterocitos, y los ácidos biliares permanecen en el lumen intestinal, pero con un paso adicional a través de los intestinos, los ácidos biliares se absorben mediante el transporte activo y pasivo.
Alrededor del 50% de los ácidos biliares se absorben en el intestino de forma pasiva. Como resultado de la escisión de los conjugados de ácidos biliares y la deshidroxilación de estos últimos por la acción de las bacterias intestinales, aumenta su solubilidad en los lípidos y se facilita la difusión pasiva.
^ Succión activa los ácidos biliares se producen exclusivamente en el íleon terminal, un fenómeno raro, conocido solo por la absorción de la vitamina B 12. Solo aquellos ácidos biliares que tienen una alta polaridad que impide su absorción pasiva, como los conjugados de taurina, están sujetos a la absorción activa. El proceso de absorción de ácidos biliares en el íleon terminal se caracteriza por signos típicos de transporte activo: cinética de saturación e inhibición competitiva. Una pequeña cantidad de ácidos biliares (7-20%) no se incluye en la absorción activa o pasiva y se elimina del cuerpo.
La presencia de ácidos biliares en el colon parece jugar un papel importante en la regulación de la consistencia de las heces. Cuando la concentración de Dioxi Ácido en el colon sobre 3 mmol / l, una cantidad significativa de electrolitos y agua se secreta en la luz intestinal, lo que conduce a la diarrea. Forma pronunciada de este "Hologennogo" diarrease puede observar con resección o enfermedad del íleon terminal, y para su tratamiento mediante la unión de ácidos biliares mediante el intercambio de iones de colestiramina.
Al ingerir los ácidos biliares absorbidos. al hígadolos conjugados se forman de nuevo, y algunos ácidos biliares secundarios experimentan hidroxilación. La pérdida de ácidos biliares en las heces (0.2-0.6 g / día) es compensada por su síntesis.
^ Piscina total de ácidos biliares. en el cuerpo hay alrededor de 3.0 g. Esta cantidad no es suficiente para asegurar la lipólisis después de una comida; En particular, el consumo de alimentos grasos requiere 5 veces más ácidos biliares. Sin embargo, el cuerpo no es deficiente en ácidos biliares, ya que circulan a través de los intestinos y el hígado muchas veces. (circulación enterohepática).La frecuencia con la que el grupo de ácidos biliares hace un ciclo completo depende del régimen dietético y varía de 4 a 12 ciclos por día.
^ Circulación de la bilirrubina. Pigmento biliar bilirrubinaal igual que los ácidos biliares y los lípidos, entra en el intestino en forma de glzhurónido. Sólo una pequeña cantidad de este compuesto polar.
Reabsorbido en la vesícula biliar y el intestino delgado. En el íleon terminal y (principalmente) en el colon, el conjugado de bilirrubina se escinde por la acción de las hidrolasas bacterianas. Al mismo tiempo la bilirrubina se convierte en urobilinógenoque, junto con otros productos de descomposición de la bilirrubina, da color marrón a las heces. Menos del 20% del urobilinógeno se absorbe, y de esta cantidad, aproximadamente el 90% vuelve a entrar en el hígado y vuelve a la bilis, y el 10% restante se excreta en la orina.
Aspectos fisiopatológicos. Los niveles elevados de urobilinógeno en la orina pueden indicar enfermedad del hígadoacompañado por una violación de la excreción de bilirrubina. La ausencia total de urobilinógeno en la orina, el color claro de las heces y la amarillez indican bloqueo completo de la vesícula biliar;en esta situación, la bilirrubina no ingresa al intestino y no se forma el urobilinógeno.
La violación más conocida y generalizada de la fisiología normal del sistema biliar es la precipitación de colesterol con la formación de cálculos biliares de colesterol. El colesterol, como la lecitina, se encuentra disuelto solo como parte de las micelas mixtas. Si la concentración aumenta xo.jecmepo.ia o bien la concentración de ácidos biliares o lecitina se encuentra por debajo de los niveles críticos,el colesterol precipita. Entre los factores que causan un aumento en el contenido de colesterol relativo están los estrógenos, una dieta con carbohidratos, sobrepeso y procesos que reducen la concentración de ácidos biliares, como la inflamación del íleon. (Enfermedad de Crohn)o resección. En algunos casos, la suplementación oral de ácidos biliares puede ser suficiente para que la bilis litogénica se convierta en alitogénica, en la cual los cálculos de colesterol pueden disolverse. Los ácidos chenodeoxicólicos y urodesoxicólicos son los más adecuados para este propósito, ya que no causan diarrea.
La manifestación clínica de alteración del metabolismo de la bilirrubina es la ictericia. La piel amarilla se asocia con niveles plasmáticos elevados de bilirrubina, que pueden ocurrir en los siguientes casos:
con aumento de la formación de bilirrubina como resultado de una mayor degradación de los eritrocitos (ictericia hemolítica);
como resultado de la interrupción del proceso de conjugación o el transporte de bilirrubina en hepatocitos, como, por ejemplo, ictericia embarazadao ictericia de maternidad de Gilbert;
con salida tardía de la bilis, por ejemplo debido a cálculos biliares o tumores localizados en la región del conducto biliar (ictericia obstructiva).