Powstanie żółci. Fizjologia powstawania żółci. Formacja żółci
Powstanie żółci zachodzi w wątrobie w wyniku czynnego i biernego transportu substancji (wody, glukozy, kreatyniny, elektrolitów, hormonów, witamin itp.) Krążących we krwi, poprzez komórki i kontakty komórka-komórka, a także aktywnej sekrecji składników żółci (kwasów żółciowych) przez hepatocyty oraz reabsorpcji wody i niektórych substancji z małych dróg żółciowych i kamieni żółciowych. Fizjologiczne znaczenie tego procesu jest różnorodne. Żółć jest uważana przede wszystkim za sekret trawienny, ponieważ kwasy żółciowe (głównie ich aniony organiczne) odgrywają kluczową rolę w wchłanianiu tłuszczu. Żółć emulguje tłuszcze, zwiększając powierzchnię, na której są hydrolizowane pod wpływem lipazy. Pod działaniem żółci jest rozpuszczanie produktów hydrolizy tłuszczu. Promuje ich wchłanianie i resyntezę triglicerydów w enterocytach. Żółć zwiększa aktywność enzymów trzustkowych i jelitowych (zwłaszcza lipaz), wzmaga hydrolizę i wchłanianie białek i węglowodanów.
Z naruszeniem trawienia tłuszczów są słabo trawione i inne substancje spożywcze, ponieważ tłuszcz otacza najmniejsze cząstki pokarmu i zapobiega działaniu na nie enzymów. W takich warunkach aktywność bakterii jelitowych prowadzi do zwiększonych procesów rozpadu, fermentacji i tworzenia się gazu.
Zawarte w układzie żółciowym i regulacyjne efekty - pobudzenie tworzenia żółci, wydalanie z żółcią, aktywność motoryczna i wydzielnicza jelita cienkiego, a także proliferacja i złuszczanie enterocytów. Żółć hamuje proces trawienia żołądka poprzez neutralizację kwasu, który dostał się do dwunastnicy i inaktywację pepsyny, przygotowując się do trawienia w jelicie. Ma również wpływ regulacyjny na aktywność ewakuacyjną kompleksu żołądkowo-dwunastniczego. Ważna jest rola żółci w wchłanianiu rozpuszczalnych w tłuszczach witamin (A, D, E i K), cholesterolu, soli wapniowych ze światła jelita.
Ponadto tworzenie się i wydzielanie żółci uważa się za swoistą metodę wydalania pewnych cząsteczek i jonów, których nie można wydalić przez nerki. Wśród nich najbardziej znaczące są cholesterol (otrzymywany jako wolny cholesterol, jego estry i kwasy żółciowe), bilirubina, a także cząsteczki miedzi i żelaza. Dlatego żółć uważana jest za płyn wydzielniczy.
Żółć składa się z 80% wody i 20% rozpuszczonych w niej substancji. Te ostatnie obejmują kwasy żółciowe i ich sole (około 65%), fosfolipidy żółciowe (około 20%, głównie z powodu lecytyny), białka (około 5%), cholesterol (4%), sprzężoną bilirubinę (0,3%), enzymy immunoglobuliny, a także liczne egzogenne i endogenne substancje wydzielane z żółcią (styreny roślinne, witaminy, hormony, leki, toksyny, jony metali - miedź, żelazo, potas, sód, wapń, cynk, magnez, rtęć, itp.). Średnio wydzielane jest 600-700 ml żółci dziennie (od 250 do 1500 ml, około 10,5 ml na 1 kg masy ciała). Jednocześnie około 500 ml / dobę całkowitej objętości zapewnia sekrecja hepatocytów i około 150 ml / dzień wydzielają się komórki dróg żółciowych.
Formacja żółci (choleresis) idzie nieprzerwanie, a przepływ żółci do dwunastnicy (cholekineza) zdarza się okresowo. Na pusty żołądek żółć wchodzi do jelita zgodnie z okresową głodową aktywnością. W czasie odpoczynku kieruje się do WB, gdzie się koncentruje, nieco zmienia swój skład i jest zdeponowany. Oprócz wody i soli wchłania się cholesterol i wolne kwasy tłuszczowe. W związku z tym rozróżnić wątrobę i żółci pęcherzyka żółciowego.
Żółć ma małą aktywność enzymatyczną; PH wątroby żółciowej wynosi 7,3-8,0. W przeciwieństwie do zawartości jelitowej nie zawiera prawie żadnych bakterii. Czynniki zapewniające sterylność żółci obejmują obecność kwasów żółciowych (działanie bakteriostatyczne), bogatą zawartość immunoglobulin, wydzielanie śluzu, względną nędzę podłoży żółciowych dla bakterii.
Żółć jest roztworem micelarnym. Cholesterol, praktycznie nierozpuszczalny w wodzie, jest transportowany w stanie rozpuszczonym w żółci ze względu na strukturę micelarną. Proces ten nazywa się rozpuszczaniem koloidalnym - solubilizacja.
Kwasy żółciowe są powierzchniowo czynnymi, amfipatycznymi (zarówno hydrofobowymi, jak i hydrofilowymi) cząsteczkami zdolnymi do samoagregacji. W tym przypadku, dzięki bardzo wąskiej szybkości koncentracji, zwanej krytycznym stężeniem micelizacji, powstają proste micele. Proste micele mają wyraźną zdolność rozpuszczania lipidów, tworząc mieszane micele.
Uważa się, że mieszane micele mają cylindryczną strukturę: cylindryczny rdzeń jest wypełniony polarnymi lipidami, a cząsteczki kwasu żółciowego są usytuowane pomiędzy polarnymi końcami cząsteczek lipidów z hydrofilowymi bokami skierowanymi w stronę środowiska wodnego, które określa ich hydrofilowość (rozpuszczalność w wodzie).
Mieszane micele zawierają niezbędne składniki - kwasy żółciowe znajdujące się na zewnątrz, fosfolipidy (głównie fosfatydylocholinę - lecytynę) i cholesterol, znajdujące się wewnątrz miceli.
Barwa żółci jest żółtawobrązowa ze względu na obecność bilirubiny, której największa część występuje w postaci bilirubiny diglukuronowej, natomiast mniejsza - w postaci bilirubiny monoglukuronidu. Związana bilirubina nie jest częścią miceli.
Ponieważ międzykomórkowe drzewo żółciowe jest przepuszczalne dla wody, pęcherzyk żółciowy i żółć wątrobowa są izotoniczne.
Formacja żółci
Wydzielanie się żółci
Kwasy żółciowe są głównym składnikiem wydzielania z żółcią, powstają tylko w wątrobie. Pierwotne kwasy żółciowe - trójhydroksycholowy (cholowy) i dihydroksycholowy (chenodeoksycholowy) syntetyzuje się w hepatocytach cholesterolowych. Wtórne kwasy żółciowe (dezoksycholowe iw małych ilościach - litocholowe) powstają w okrężnicy z pierwotnej w wyniku modyfikacji bakteryjnej nuklearnych grup hydroksylowych (7a-dehydroksylacja pod kontrolą bakterii beztlenowych). Trzeciorzędowe kwasy żółciowe (głównie ursodeoksycholik) powstają w wątrobie przez izomeryzację wtórnych kwasów żółciowych.
Synteza kwasów żółciowych z cholesterolu jest regulowana przez mechanizm ujemnego sprzężenia zwrotnego: zmniejszony powrót kwasów żółciowych do hepatocytów z krwią wrotną prowadzi do zwiększenia ich biosyntezy; każdemu wzrostowi syntezy kwasów żółciowych towarzyszy odpowiedni wzrost tworzenia cholesterolu.
Kwasy żółciowe są sprzężone w wątrobie z aminokwasami glicyny (około 80%) lub tauryny (około 20%). Po spożyciu pokarmów bogatych w węglowodany, ilość kwasów glikocholowych wzrasta w żółci, aw dietach wysokobiałkowych - w taurocholii. Wiązanie aminokwasów zapobiega wchłanianiu kwasów żółciowych w drogach żółciowych i początkowych odcinkach jelita cienkiego (tylko w końcowym jelicie krętym iw jelicie grubym). Pod wpływem bakterii możliwa jest hydroliza soli żółciowych z powstawaniem kwasów żółciowych, glicyny lub tauryny.
Po biosyntezie kwasów żółciowych, jak zauważono powyżej, ich grupa karboksylowa jest związana z grupą aminową glicyny lub tauryny. Z chemicznego punktu widzenia takie wiązanie przekształca słaby kwas w silniejszy. Związane kwasy żółciowe są najbardziej rozpuszczalne przy niskim pH i bardziej odporne na strącanie z jonami wapnia (Ca 2+) niż niezwiązane kwasy żółciowe. Z biologicznego punktu widzenia proces wiązania powoduje, że kwasy żółciowe nie są w stanie przeniknąć przez błony komórkowe. Dlatego są one absorbowane w drogach żółciowych lub jelicie cienkim, albo w obecności cząsteczek nośnikowych, albo jeśli kwasy żółciowe ulegają rozkładowi bakteryjnemu. Jedynym wyjątkiem od tej zasady są kwasy żółciowe związane z dihydroksylem glicyny, ponieważ mogą one być wchłaniane pasywnie, jeśli otrzymują jon wodorowy (H +).
Większość kwasów żółciowych (około 85-90%), otrzymanych wraz z przepływem żółci do jelita cienkiego, jest absorbowana do krwi. Jednak większość powiązanych kwasów żółciowych, które są wydzielane do jelita cienkiego, są wchłaniane w stanie nienaruszonym. Mniejsza część kwasu nie jest wchłaniana w stanie nienaruszonym, ponieważ ulega rozkładowi bakteryjnemu w dystalnej części jelita cienkiego. Są one wchłaniane pasywnie, a po powrocie do wątroby są ponownie związane i wydzielane do żółci. Pozostałe 10-15% kwasów żółciowych wydalanych jest z organizmu głównie z kałem. Ta utrata kwasów żółciowych jest kompensowana przez ich syntezę w hepatocytach.
Ten proces rozszczepiania jelita i reabsorpcji w wątrobie jest normalną częścią metabolizmu kwasów żółciowych. Mała część kwasów żółciowych, nie wchłonięta, wchodzi do okrężnicy. Tutaj kończy się proces dzielenia. Ponadto, wtórne kwasy żółciowe powstają w okrężnicy pod wpływem bakterii beztlenowych (patrz powyżej).
Kwasy deoksycholowy i litocholowy są częściowo wchłaniane w okrężnicy i ponownie wchodzą do wątroby. Po powrocie metabolizm tych kwasów żółciowych jest inny. Kwas dezoksycholowy wiąże się z glicyną lub tauryną i krąży z pierwszorzędowymi kwasami żółciowymi. Należy zauważyć, że u osób w podeszłym wieku kwas dezoksycholowy jest głównym kwasem żółciowym w kompozycji żółci. Kwas litocholowy nie tylko wiąże się z glicyną lub tauryną, ale dodatkowo siarczanami w pozycji C-3. Takie "podwójne" wiązanie zmniejsza możliwość wchłaniania w jelitach, w wyniku czego kwas litocholowy jest szybko tracony z kompozycji krążących kwasów żółciowych - jego zawartość w żółci rzadko przekracza 5%. Większość (95%) kwasów żółciowych, które są częścią żółci dorosłych, to kwasy chololowe, szenodoksycholowe i dezoksycholowe.
Proces wiązania kwasów żółciowych w hepatocytach jest bardzo skuteczny, więc żółciowe kwasy żółciowe są całkowicie obecne w postaci skoniugowanej. Późniejsze cięcie bakteryjne i dehydroksylacja powodują, że kwasy żółciowe w kale znajdują się w stanie niezwiązanym.
Opisany powyżej proces dehydroksylowania pierwotnych kwasów żółciowych zmniejsza ich zdolność do rozpuszczania w wodzie. Tworzenie się wtórnych kwasów żółciowych u ludzi nie ma praktycznie żadnego znaczenia fizjologicznego, w przeciwieństwie do zwierząt. Nadmierna absorpcja kwasu dezoksycholowego w okrężnicy zwiększa ryzyko powstania kamieni żółciowych cholesterolu. Kwas litocholowy uważany jest za hepatotoksyczny. W doświadczeniach na zwierzętach wykazano, że akumulacja kwasu litocholowego w wątrobie prowadzi do jego porażki. Jednak nie udowodniono jeszcze, że zwiększenie wchłaniania tego kwasu u ludzi prowadzi do zaburzeń czynności wątroby.
Kwas ursodeoksycholowy, a także kwas dezoksycholowy wiążą się w wątrobie i cyrkulują z pierwszorzędowymi kwasami żółciowymi. Jednak droga metaboliczna tego kwasu jest bardzo krótka, a zawartość związanego kwasu ursodeoksycholowego w żółci nigdy nie przekracza 5% całkowitej ilości kwasów żółciowych. Uważa się, że tworzenie kwasu ursodeoksycholowego nie ma istotnego znaczenia fizjologicznego.
Kwasy żółciowe są silnymi rozpuszczalnikami tłuszczów, dlatego są cytotoksyczne w stężeniach, które osiągają krytyczne stężenie micelizacji. Jednocześnie, związane kwasy chenodeoksycholowe i deoksycholowe obecne w żółci są bardziej toksyczne niż te związane z cholem i ursodeoksycholem. Chociaż kwasy żółciowe wykazują demonstracyjnie cytotoksyczność in vitro, nabłonek przewodów żółciowych i jelita cienkiego nigdy nie ulega uszkodzeniu w wyniku ich wysokiego stężenia. Jest to związane, po pierwsze, z obecnością innych lipidów (fosfolipidów w żółci i kwasów tłuszczowych w jelicie), które zmniejszają monometryczne stężenie kwasów żółciowych, a po drugie, z obecnością glikolipidów i cholesterolu w apikalnych błonach komórek nabłonkowych, które zwiększają odporność komórek nabłonkowych na uszkodzenia. działanie anionów kwasu żółciowego.
Kwasy cholowy, chenodeoksycholowy i deoksycholowy są wchłaniane i podlegają krążeniu jelitowo-wątrobowemu do 6-10 razy dziennie. Kwas litocholowy jest słabo wchłaniany, a jego ilość w żółci jest niewielka. Pula kwasów żółciowych wynosi zwykle około 2,5 g, a codzienne wytwarzanie pierwotnych kwasów żółciowych, chol i chenodeoksycholi wynosi średnio około 330 i 280 mg, odpowiednio.
Regulacja powstawania żółci
Jak wiadomo, powstawanie żółci odbywa się w sposób ciągły, ale intensywność tego procesu jest różna. Zwiększyć wydalanie żółci i przyjmowanie pokarmów. Odruchowe działanie na cholerezy występuje podczas stymulacji receptorów przewodu żołądkowo-jelitowego (GIT), narządów wewnętrznych i uwarunkowanych efektów odruchowych.
Regulację wegetatywną zapewnia cholinergiczny układ przywspółczulny (zwiększenie powstawania żółci) i współczulne włókna nerwu adrenergicznego (zmniejszenie powstawania żółci).
Regulacja humoralna jest prowadzona przez samą żółć z powodu obecności krążenia jelitowo-wątrobowego kwasów żółciowych i mechanizmu ujemnego sprzężenia zwrotnego (patrz powyżej). Sekretina stymuluje wydzielanie żółci, wydzielanie wody i elektrolitów w jej składzie. Glukagon, gastryna i cholecystokinina mają słabsze działanie stymulujące.
Wydalanie z żółcią
Wydzielanie żółci uważa się za swoisty sposób usuwania cząsteczek i jonów z organizmu, których nie można wydalić przez nerki. Najważniejsze z nich to cholesterol (jako taki i w postaci kwasów żółciowych) i bilirubina, a także jony miedzi, żelaza itp.
Główne składniki wydalania z żółcią
Cholesterol, praktycznie nierozpuszczalny w wodzie, jest transportowany w kompozycji mieszanych miceli utworzonych, jak wspomniano powyżej, przez kwasy żółciowe, fosfolipidy i sam cholesterol.
Cząsteczki fosfolipidów spełniają dwie ważne funkcje. Po pierwsze, znacznie zwiększają micelarną rozpuszczalność cholesterolu, ponieważ mieszane micele zawierające fosfolipidy rozpuszczają znacznie więcej cholesterolu niż proste micele zawierające tylko cząsteczki kwasu żółciowego. Po drugie, obecność fosfolipidów w żółci zmniejsza krytyczne stężenie micelizacji i monometryczne stężenie kwasów żółciowych. W wyniku tego zmniejsza się aktywność powierzchniowa i cytotoksyczność wątrobowej żółci.
Wraz z przepływem żółci, mieszane micele wchodzą do jelita cienkiego, gdzie dalsza transformacja ich składników jest inna. Kwasy żółciowe rozpuszczają lipidy, zapewniając ich wchłanianie i są absorbowane w bardziej odległych częściach jelita. Żółciowe fosfolipidy, nierozpuszczalne w wodzie, hydrolizują w jelitach i nie biorą udziału w krążeniu jelitowo-wątrobowym. Kwasy żółciowe regulują ich wydalanie i stymulują syntezę.
Ponieważ około 1/3 cholesterolu jest wchłaniane, 2/3 jego ilości jest wydalane. U osoby dorosłej równowaga cholesterolu jest zapewniona przez jego uwalnianie jako takie (około 600 mg / dzień) lub w postaci kwasów żółciowych (około 400 mg / dzień). Jednocześnie krążenie jelitowo-wątrobowe kwasów żółciowych uważa się za opóźniony sposób wydalania cholesterolu.
Należy zauważyć, że w porównaniu ze zwierzętami człowiek ma wyższy udział cholesterolu w żółci. Uważa się, że jest to wynikiem wadliwej konwersji cholesterolu do kwasów żółciowych, a także stosunkowo niskiej szybkości wydzielania kwasów żółciowych. Dlatego żółć jest przesycona cholesterolem u około 25% osób w podeszłym wieku, a w 10-15% populacji tworzą się kamienie cholesterolu (kamica żółciowa).
Jak zauważono powyżej, synteza kwasów żółciowych z cholesterolu jest regulowana przez mechanizm ujemnego sprzężenia zwrotnego.
Żółć jest głównym sposobem usuwania bilirubiny z organizmu. Bilirubina - Produkt końcowy rozkładu hemu, w jego strukturze chemicznej jest tetrapirol. Największa ilość bilirubiny (80-85%) pochodzi z rozkładającej się hemoglobiny starzejących się czerwonych krwinek i przedwcześnie zwijających się nowo powstałych czerwonych krwinek w szpiku kostnym lub w łóżku krążącym (tak zwana nieskuteczna erytropoeza). Pozostała część bilirubiny powstaje w wyniku zniszczenia innych białek zawierających hem (na przykład cytochromu P-450 itd.) W wątrobie, a znacznie mniej w tkance pozawątrobowej. Należy zauważyć, że katabolizm hemoglobiny erytrocytów w bilirubinę występuje przede wszystkim w makrofagach śledziony, wątroby i szpiku kostnego.
Źródła tworzenia bilirubiny (w procentach)

Jak wiadomo, nieskoniugowana bilirubina jest hydrofobowa (nierozpuszczalna w wodzie) i potencjalnie toksyczna substancja, która krąży w osoczu w stanie blisko spokrewnionym z albuminą i nie może być wydalana z moczem. Zdolność organizmu do usuwania bilirubiny wiąże się z usunięciem tego ostatniego z osocza krwi przez komórkę wątroby, następnie koniugacja z kwasem glukuronowym i uwolnienie do żółci już rozpuszczalnego w wodzie związku (związanego lub bezpośredniego bilirubiny). Proces koniugacji przebiega poprzez enzym mikrosomalny glukuronylotransferazę difosforanu urydyny. Koniugacja bilirubiny z kwasem glukuronowym ma najważniejsze znaczenie fizjologiczne, jednak koniugacja bilirubiny z siarczanami, glukozą i ksylozą występuje w niewielkich ilościach.
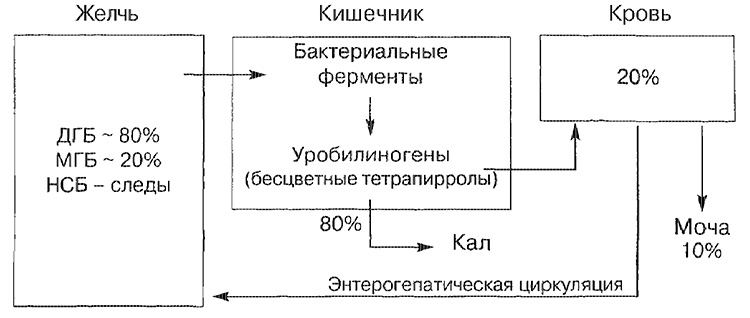
Główne szlaki transportowe dla bilirubiny
NSB - niezwiązana bilirubina; MGB - monoglukuronid bilirubiny; DGB - bilirubina diglukuronidu.
Wydzielanie sprzężonej bilirubiny do kanalików żółciowych następuje z udziałem rodziny zależnych od ATP wieloswoistych białek transportowych dla anionów organicznych.
Większość bilirubiny bilirubiny (około 80%) jest reprezentowana w postaci bilirubiny diglukuronidowej, mniejszej części - w postaci monoglukuronidu, a tylko niewielka jej ilość jest reprezentowana przez niezwiązaną postać. Bilirubina, która dostała się do jelita, jest rozkładana w końcowej części jelita cienkiego iw jelicie grubym przez enzymy bakteryjne (beta-glukuronidazę), która przekształca się w bezbarwny tetrapirol (urobilinogen). Około 20% utworzonych urobilinogenów jest resorbowanych i koniecznie wydalanych z moczem, a także z żółcią (krążenie krążeniowo-jelitowe).
Większość autorów zgadza się, że normalne stężenie bilirubiny całkowitej w surowicy, określone przez reakcję diazo Van Der Bergha, zwykle nie przekracza 1 mg% (0,3-1 mg%, lub 5-17 μmol / l). Tylko mniej niż 5% bilirubiny jest prezentowane w postaci związanej. Zwiększenie poziomu bilirubiny we krwi (hiperbilirubinemia) i jej nagromadzenie w tkankach prowadzi do pojawienia się żółtaczki, która z reguły staje się zauważalna przy wartościach przekraczających 2,5-3 mg%.
Jak wspomniano powyżej, bilirubina krąży we krwi w stanie związanym z białkami i prawie nie penetruje płynów tkankowych o niskiej zawartości białka. Pod tym względem wysięki są bardziej żółte niż przesięki. Bilirubina wiąże się dobrze z tkanką elastyczną, co wyjaśnia wczesne żółte zabarwienie twardówki, skóry i ścian naczyń krwionośnych w hiperbilirubinemii. Przedłużająca się cholestaza powoduje pojawienie się zielonkawego odcienia skóry, co tłumaczy się depozycją biliverdinu.
Należy podkreślić, że ciśnienie w przewodach żółciowych, na którym występuje wydzielanie żółci, zwykle wynosi 150-200 mm wody. Art. Jego wzrost do 350 mm wody. Art. hamuje wydzielanie żółci, co prowadzi do rozwoju żółtaczki. W przypadku całkowitego zaprzestania wydzielania bilirubiny i kwasów żółciowych, żółć ulega odbarwieniu (tzw. Biała żółć).
Ponadto, żółć jest sposobem wydalania tłuszcze roślinne, leki lipofilowe i ich metabolity, różne ksenobiotyki obecne w roślinach, lipofilowe metabolity, witaminy rozpuszczalne w tłuszczach i hormony steroidowe.
Bilans żelaza i miedzi ciało jest również wspomagane przez wydalanie żółciowe tych metali. Oba kationy są wydzielane do żółci przez stymulowaną ATP pompę kanałową. Z żółcią występuje i uwalnianie innych metali.
Żółć
Prąd żółciowy w układzie żółciowym jest spowodowany różnicą ciśnienia w różnych jego sekcjach i dwunastnicy, tonem zwieracza, skurczem włókien mięśni gładkich w tunelu czasoprzestrzennym i kanałach (pierwsze pojedyncze włókna mięśniowe pojawiają się w wewnątrzwątrobowych drogach żółciowych położonych w bezpośrednim sąsiedztwie bramek wątroby). Aktywność całego systemu jest zwykle dobrze skoordynowana i kontrolowana przez mechanizmy nerwowe i humoralne.
W kamieniach żółciowych stężenia żółci są zdeponowane i zwiększone z 10-krotnym wzrostem stężenia lipidów, soli kwasów żółciowych, żółciowych pigmentów itp. W odpowiednim momencie jest uwalniany do OZhP i dalej do dwunastnicy. Przejście współrzędnych zoladka aparat zwieracza. Poza procesem trawienia w dwunastnicy zwieracz fiolki jest zamknięty. W tym czasie RI i torbielowaty przewód są rozluźnione, co pozwala żółci wyprodukowanej przez wątrobę wejść do światła RI. Błona śluzowa ZH pochłania wodę, jony. W tym samym czasie żółć staje się bardziej skoncentrowana. Produkty śluzu pozwalają żółci być w stanie koloidalnym.
Ciśnienie w świetle gruczołu w spoczynku jest znacznie mniejsze niż w przewodach żółciowych i wynosi 60-185 mm wody. Art. Różnica w ciśnieniu jest fizjologiczną podstawą przepływu żółci w pęcherzyku żółciowym z zamkniętym zwieraczem Oddi. W procesie trawienia ze względu na zmniejszenie ciśnienia GF wzrasta do 150-260 mm wody. st, zapewniając przepływ żółci do dwunastnicy przez luźne ampułki zwieracza. Kiedy żółć zaczyna wpływać do dwunastnicy, ciśnienie w przewodach stopniowo zmniejsza się (przy każdym posiłku zawartość tłuszczu zmniejsza się 1-2 razy).
Okres pierwotnej reakcji układu żółciowego, ze względu na rodzaj, zapach pokarmu i jego odbiór, trwa około 7-10 minut. Następnie następuje okres ewakuacji (główny lub okres opróżniania ZHP), podczas którego, na tle przemian skurczów i relaksacji ZP, pęcherzyk żółciowy, a następnie żółć wątrobowa wypływają do dwunastnicy.
Odruchowa stymulacja cholekineza (warunkowo i bezwarunkowo-odruchowa) występuje z powodu receptorów zlokalizowanych w jamie ustnej, żołądku i dwunastnicy, poprzez nerwy błędne. Regulacja humoralna odbywa się głównie pod wpływem cholecystokininy, która ma działanie stymulujące na GF, powodując jej zmniejszenie. Ponadto, słabe cięcia w GI występują pod wpływem gastryny, sekretyny i Bombezinu. Przeciwnie, glukagon, kalcytonina, antycholecystokinina, peptyd naczyniowo-jonowy i polipeptyd trzustkowy hamują redukcję GI.
Żółtka, tłuszcze, mleko i mięso są silnymi środkami pobudzającymi wydzielanie żółci.
Zaburzenia aparatu zwieracza i ruchliwość gonad mogą powodować zmiany w kierunku i szybkości przepływu żółci, dyskinezę, refluks wydzielin trzustkowych i zawartość dwunastnicy w ACP oraz żółć w przewodzie trzustkowym, która jest podstawą do rozwoju wielu stanów patologicznych i chorób.
Sole żółciowe, sprzężona bilirubina, cholesterol, fosfolipidy, białka, elektrolity i woda są wydzielane przez hepatocyty do kanalików żółciowych. Aparat wydzielania żółci zawiera białka transportowe membrana rurkowa, wewnątrzkomórkowe organellei struktury cytoszkielet. Szczelne kontaktypomiędzy hepatocytami światło kanalików jest oddzielone od układu krążenia w wątrobie.
Rurowa membrana zawiera białka transportowe dla kwasów żółciowych, bilirubiny, kationów i anionów. Microvilli zwiększa powierzchnię. Organelle są reprezentowane przez aparat Golgiego i lizosomy. Za pomocą pęcherzyków białka (np. IgA) są transportowane z błony sinusoidalnej do błony kanałowej, transport białek transportowych zsyntetyzowanych w komórce w celu uzyskania cholesterolu, fosfolipidów i ewentualnie kwasów żółciowych z mikrosomów do błony rurkowej.
Cytoplazma hepatocytów wokół kanalików zawiera strukturę cytoszkieletu: mikrotubule, mikrofilamentyi pośrednie włókna ciągłe.
Mikrotubule powstają w wyniku polimeryzacji tubuliny i tworzą sieć w komórce, szczególnie w pobliżu membrany podstawno-bocznej i aparatu Golgiego, biorącego udział w transporcie pęcherzykowym, w którym pośredniczy receptor, wydzielaniu lipidów i pod pewnymi warunkami - kwasy żółciowe. Tworzenie się mikrotubuli jest hamowane przez kolchicynę.
Oddziaływanie spolimeryzowanych (F) i wolnych (G) aktyny bierze udział w budowie mikrowłókien. Mikrofilamenty, koncentrujące się wokół rurowej membrany, określają kurczliwość i ruchliwość kanalików. Phalloidin, która wzmaga polimeryzację aktyny i cytochalasynę B, która ją osłabia, hamuje ruchliwość kanalików i powoduje cholestazę.
Włókna pośrednie składają się z cytokeratyny i tworzą sieć między błonami plazmatycznymi, jądrem, organellami wewnątrzkomórkowymi i innymi strukturami cytoszkieletu. Zerwanie pośrednich włókien ciągłych prowadzi do przerwania wewnątrzkomórkowych procesów transportu i zamazania światła kanalików.
Woda i elektrolity wpływają na skład cewkowego wydzielania, przenikając przez ścisłe kontakty między hepatocytami z powodu gradientu osmotycznego między światłem kanalikowym a przestrzeniami Disse. (prąd paracellularny).Integralność ścisłych kontaktów zależy od obecności na wewnętrznej powierzchni błony komórkowej białka ZO-1 o masie cząsteczkowej 225 kDa. Zerwaniu ścisłych kontaktów towarzyszy wejście rozpuszczonych większych cząsteczek do kanalików, co prowadzi do utraty gradientu osmotycznego i rozwoju cholestazy. Jednocześnie można zaobserwować regurgitację kanalikowej żółci w sinusoidzie.
Kanały żółciowe wpływają do przewodów, czasami nazywane cholangiolami lub kanałami Goeringa. Ductulas są zlokalizowane głównie w obszarach portalowych i wpływają do międzyzbłonkowych dróg żółciowych, które są pierwszymi z dróg żółciowych, którym towarzyszą odgałęzienia tętnicy wątrobowej i żyły wrotnej i znajdują się w składzie triad portalowych. Kanały między warstwowe, łączące się, tworzą kanały przegrodowe, aż do pojawienia się dwóch głównych przewodów wątrobowych, wyłaniających się z prawego i lewego płata w szczelinie wrotnej wątroby.
Wydzielanie żółci
Tworzenie żółci odbywa się przy udziale wielu niestabilnych procesów transportowych. Jego wydzielanie jest względnie niezależne od ciśnienia perfuzji. Całkowity prąd żółci u ludzi wynosi około 600 ml / dobę. Hepatocyty zapewniają wydzielanie dwóch frakcji żółci: zależą od kwasów żółciowych ("225 ml / dzień) i nie zależą od nich (" 225 ml / dzień). Pozostałe 150 ml / dzień są wydzielane przez komórki dróg żółciowych.
Wydzielanie soli kwasów żółciowych jest najważniejszym czynnikiem w powstawaniu żółci (frakcja zależna od kwasów żółciowych).Woda porusza się w wyniku osmotycznie czynnych soli kwasów żółciowych. Zmiany aktywności osmotycznej mogą regulować przepływ wody do żółci. Istnieje wyraźna korelacja pomiędzy wydzielaniem soli kwasów żółciowych a przepływem żółci.
Istnienie frakcji żółciowej, która nie zależy od kwasów żółciowych, udowadnia możliwość powstania żółci, która nie zawiera soli kwasów żółciowych. W ten sposób możliwe jest kontynuowanie przepływu żółci, pomimo braku wydzielania soli kwasów żółciowych; wydzielanie wody w tym przypadku jest spowodowane innymi osmotycznie czynnymi substancjami rozpuszczalnymi, takimi jak glutation i wodorowęglany.
Komórkowe mechanizmy wydzielania żółci
Hepatocyt to polarna sekrecyjna komórka nabłonka mająca boki podstawno-boczne (boczne i boczne) i wierzchołkowe (rurkowe).
Tworzenie się żółci obejmuje wychwytywanie kwasów żółciowych i innych jonów organicznych i nieorganicznych, ich transport przez błonę podstawno-boczną (sinusoidalną), cytoplazmę i błonę kanalikową. Procesowi temu towarzyszy filtracja osmotyczna wody zawartej w hepatocytach i przestrzeni okołokomórkowej. Identyfikacja i charakterystyka białek transportowych membran sinusoidalnych i rurkowych są złożone. Szczególnie trudne jest badanie aparatury wydzielniczej kanalików, jednak do tej pory opracowano i udowodniono, że technika otrzymywania podwojonych hepatocytów w hodowli krótkotrwałej jest wiarygodna w wielu badaniach: klonowanie białek transportowych pozwala nam scharakteryzować funkcję każdego z nich osobno.
Proces powstawania żółci zależy od "obecności pewnych białek nośnikowych w błonie podstawno-bocznej i rurowej, Na +, K pełni rolę siły napędowej sekrecji + - ATPaza błony podstawno-bocznej, zapewniająca gradient chemiczny i potencjalną różnicę między hepatocytem a otaczającą przestrzenią. Na +, K + - ATPase wymienia trzy wewnątrzkomórkowe jony sodu dla dwóch zewnątrzkomórkowych jonów potasowych, utrzymując gradient stężenia sodu (wysoki na zewnątrz, niski wewnątrz) i potas (niski na zewnątrz, wysoki wewnątrz). W rezultacie zawartość komórek ma ładunek ujemny (-35 mV) w porównaniu do przestrzeni pozakomórkowej, co ułatwia wychwytywanie dodatnio naładowanych jonów i wydalanie ujemnie naładowanych jonów. Na +, K + -ATPazy nie wykrywa się w błonie rurkowej. Przepływ błon może wpływać na aktywność enzymu.
Przechwyć na powierzchni membrany sinusoidalnej
Membrana podstawno-boczna (sinusoidalna) ma wiele systemów transportowych do wychwytywania anionów organicznych, których specyficzność substratowa częściowo się pokrywa. Charakterystykę białek nośnikowych podano wcześniej na podstawie badania komórek zwierzęcych. Ostatnie klonowanie ludzkich białek transportowych pozwoliło lepiej scharakteryzować ich funkcję. Białko transportowe dla anionów organicznych (białka transportującego aniony organiczne - OATP) jest niezależne od sodu, niesie cząsteczki wielu związków, w tym kwasów żółciowych, bromosulfaleiny i prawdopodobnie bilirubiny. Uważa się, że transport bilirubiny w hepatocytach jest również przeprowadzany przez innych przewoźników. Wychwytywanie kwasów żółciowych sprzężonych z tauryną (lub glicyną) przeprowadza się za pomocą białka transportującego sód / taurocholan (białko transportujące sód / kwas żółciowy - NTCP).
Transfer jonów przez membranę podstawno-boczną obejmuje białko, które wymienia Na + / H + i reguluje pH wewnątrz komórki. Ta funkcja jest również wykonywana przez białko kotransportowe dla Na + / HCO 3 -. Na powierzchni błony podstawno-bocznej wychwytywane są także siarczany, niezestryfikowane kwasy tłuszczowe i organiczne kationy.
Transport wewnątrzkomórkowy
Transport kwasów żółciowych w hepatocytach odbywa się przy użyciu białek cytozolowych, wśród których główną rolę odgrywa dehydrogenaza Z-hydroksysteroidowa. S-transferaza glutationowa i białka wiążące kwasy tłuszczowe mają mniejsze znaczenie. Retikulum endoplazmatyczne i aparat Golgiego są zaangażowane w przenoszenie kwasów żółciowych. Wydaje się, że transport pęcherzykowy jest aktywowany tylko wtedy, gdy kwasy żółciowe są znacząco wstrzyknięte do komórki (w stężeniach przekraczających fizjologiczne).
Transport białek w fazie ciekłej i ligandów, takich jak IgA i lipoproteiny o niskiej gęstości, odbywa się przez pęcherzykową transcytozę. Czas transferu od boczno-bocznego do błony rurkowej wynosi około 10 minut. Mechanizm ten odpowiada tylko za niewielką część całkowitego prądu żółci i zależy od stanu mikrotubul.
Wydzielanie cewkowe
Rurowa membrana jest wyspecjalizowaną częścią błony komórkowej hepatocytów zawierającą białka transportowe (głównie zależne od ATP) odpowiedzialne za przenoszenie cząsteczek do żółci na gradient stężenia. Enzymy, takie jak fosfataza alkaliczna, GGTP są również zlokalizowane w błonie kanałowej. Przenoszenie glukuronidów i koniugatów glutationu-S (na przykład diglukuronidu bilirubiny) prowadzi się za pomocą kanału wieloswoistego białka transportowego dla anionów organicznych (gruczołu cMOAT); transporter - SWAT), którego funkcja jest częściowo kontrolowana przez ujemny potencjał wewnątrzkomórkowy. Prąd żółci, który nie zależy od kwasów żółciowych, jest najwyraźniej określony przez transport glutacji, a także kanalikowe wydzielanie wodorowęglanu, ewentualnie przy udziale białka wymieniającego Cl - / HCO 3 -.
Ważna rola w transporcie substancji przez błonę kanalikową należy do dwóch enzymów z rodziny glikoprotein P; oba enzymy są zależne od ATP. Białko 1 oporności wielolekowej 1 (MDR1) oporności wielolekowej 1 przenosi kationy organiczne, a także usuwa leki cytostatyczne z komórek nowotworowych, powodując ich oporność na chemioterapię (stąd nazwa białko). Endogenny substrat MDR1 jest nieznany. MDR3 transportuje fosfolipidy i działa jako flippaza dla fosfatydylocholiny. Funkcja MDR3 i jej znaczenie dla sekrecji fosfolipidów w żółci są udoskonalane w doświadczeniach na myszach, które nie mają glikoproteiny mdr2-P (analogicznie do ludzkiego MDR3). W przypadku braku fosfolipidów w żółci, kwasy żółciowe powodują uszkodzenie nabłonka dróg żółciowych, zapalenie przewodów i zwłóknienie okołopanialne.
Woda i nieorganiczne jony (zwłaszcza sód) są wydalane do kapilar żółciowych wzdłuż gradientu osmotycznego poprzez dyfuzję przez ujemnie naładowane półprzepuszczalne ciasne kontakty.
Wydzielanie żółci jest regulowane przez wiele hormonów i wtórnych przekaźników, w tym cAMP i kinazę białkową C. Zwiększenie stężenia wewnątrzkomórkowego wapnia hamuje wydzielanie żółci. Przejście żółci przez kanaliki jest spowodowane mikrofilamentami, które zapewniają ruchliwość i kurczenie się kanalików.
Cieczne wydzielanie
Komórki nabłonkowe kanałów dystalnych wytwarzają tajemnicę bogatą w wodorowęglany, która modyfikuje skład rurkowej żółci (tzw. prąd przewodzący, żółć).Proces sekrecji wytwarza cAMP, niektóre białka transportujące błonę, w tym białko, które wymienia Cl - / HCO 3 - i regulator przewodzenia transmembranowego w mukowiscydozie -kanał membranowy dla Cl -, regulowany cAMP. Wydzielanie kanału jest stymulowane przez sekretynę.
Zakłada się, że kwas ursodeoksycholowy jest aktywnie absorbowany przez komórki przewodowe, zamieniany na wodorowęglany, zawracany do wątroby, a następnie wydalany ponownie do żółci ("bocznik trójdzielno-wątrobowy"). Być może wyjaśnia to efekt żółciopędny kwasu ursodeoksycholowego, któremu towarzyszy wysokie wydzielanie wodorowęglanu żółciowego w eksperymentalnej marskości wątroby.
Ciśnienie w drogach żółciowych, przy którym występuje wydzielanie żółci, wynosi zwykle 15-25 cm wody. Art. Zwiększyć ciśnienie do 35 cm wody. Art. prowadzi do zahamowania wydzielania żółci, rozwoju żółtaczki. Wydzielanie bilirubiny i kwasów żółciowych można całkowicie zatrzymać, podczas gdy żółć staje się bezbarwna (biały żółć)i przypomina płyn śluzowy.
Żółć to izosmotyczna ciecz w osoczu składająca się z wody, elektrolitów i substancji organicznych (kwasów żółciowych, fosfolipidów, cholesterolu, bilirubiny). Kwasy żółciowe (lub ich sole) są głównym organicznym składnikiem żółci. Kwasy żółciowe przedostają się do żółci z dwóch źródeł: (1) pierwotnych kwasów żółciowych (cholek i chenodeoksycholików), które są syntetyzowane z cholesterolu w wątrobie; (2) wtórne kwasy żółciowe (dezoksycholowe, litocholowe i ursodeoksycholowe) powstają w wyniku działania bakterii jelitowych z pierwotnych kwasów żółciowych. Same kwasy żółciowe składają się z dwóch ważnych składników, które określają ich właściwości fizjologiczne i fizykochemiczne: (1) rdzeń steroidowy z podstawnikami hydroksylowymi; (2) alifatyczny łańcuch boczny (zdjęcie 7-4).
Ryc. 7-4.
Kwasy żółciowe składają się z dwóch składników - jądra z hydroksylowymi końcami i alifatycznym łańcuchem bocznym. Na figurze pokazano kwas cholowy jako przykład 3-hydroksykwasu (3 (3 -, 7 (3 -, 12 (3-OH). Innymi przykładami są kwasy żółciowe zawierające dezoksycholan (3a -, 12a-OH), chenodeoksycholan (3a -, 7a-OH) i litocholan (3a-OH).
U większości ssaków pierwotne kwasy żółciowe zawierają od trzech do siedmiu podstawników hydroksylowych, których liczba wpływa na ich rozpuszczalność w wodzie (hydrofilowość). Krótko po utworzeniu pierwotne kwasy żółciowe ulegają modyfikacji w końcowej grupie karboksylowej. Dzieje się tak w fazie wątrobowej krążenia jelitowo-wątrobowego wtórnych kwasów żółciowych i ich koniugacji z glicyną lub tauryną. Obecność hydrofilowych (składników hydroksylowych i wiązań amidowych alifatycznego łańcucha bocznego) i składników hydrofobowych (rdzeń steroidowy) umożliwia sprzężonym cząsteczkom kwasu żółciowego działanie jako związek amfoteryczny. To daje im możliwość tworzenia miceli (agregatów polimolekularnych) powyżej krytycznego stężenia micelarnego. Z kolei cząsteczki kwasów żółciowych mogą rozpuszczać inne substancje amfoteryczne (cholesterol, fosfolipidy) z tworzeniem się mieszanych miceli. Ta detergentowa rola kwasów żółciowych jest ważna dla stabilizacji stanu fizyczno-chemicznego żółci, trawienia i wchłaniania tłuszczów.
Synteza kwasów żółciowych z cholesterolu jest regulowana przez mechanizm negatywnego sprzężenia zwrotnego, chociaż natura regulacji na poziomie molekularnym i biochemicznym nie jest jeszcze w pełni zrozumiała. Cykloksylacja 7-hydroksylowa mikrosomów jest kluczowym etapem w syntezie kwasów żółciowych. Kwas Chenodeoksycholowy, stosowany do rozpuszczania kamieni pęcherzyka żółciowego, hamuje syntezę kwasów żółciowych, a tym samym podnosi poziom cholesterolu we krwi. Stosując kwas ursodeoksycholowy, takich zmian nie obserwuje się nawet przy długotrwałym leczeniu.
Tworzenie się żółci odbywa się zarówno na powierzchniach sinusoidalnych, jak i cylindrycznych błony hepatocytów i jest procesem wewnątrzkomórkowym i parakomórkowym. W przeciwieństwie do filtracji kłębuszkowej w nerkach, które biernie pod wpływem sił hydrostatycznych, podczas formowania się żółci, następuje czynne przeniesienie składników organicznych i nieorganicznych do światła kanalików i pasywny transport wody. Tak więc procesy wydzielania żółci są podobne do procesów sekrecji w acini trzustki, nabłonku kanalików nerkowych. Tworzenie kanalików żółciowych można podzielić na dwa typy (ryc. 7-5): (1) powstawanie żółci, w zależności od wydzielania kwasów żółciowych, zdefiniowane jako stosunek ilości żółci wydzielanej do kanalików do ilości wydzielanych soli żółciowych; (2) powstawanie żółci, niezależne od wydzielania kwasu żółciowego, które może być reprezentowane jako aktywna sekrecja nieorganicznych elektrolitów i innych substancji i jest odzwierciedlone na wykresie jako przecięcie y tej linii. Innymi słowy, tworzenie żółci w związku z wydzielaniem kwasu jest natężeniem przepływu żółci, w zależności od obecności osmotycznie czynnych soli żółciowych w drogach żółciowych i tworzenia żółci niezwiązanej z wydzielaniem kwasu pod nieobecność soli żółciowych. Stosunek szybkości tworzenia żółci i tworzenia soli kwasów żółciowych jest nieliniowy z małymi ilościami wydzielanej żółci i nie może odpowiadać zależności liniowej pokazanej na ryc. 7-5. Dlatego oba typy formowania się żółci powinny być uważane za wzajemnie powiązane wskaźniki powstawania żółci.
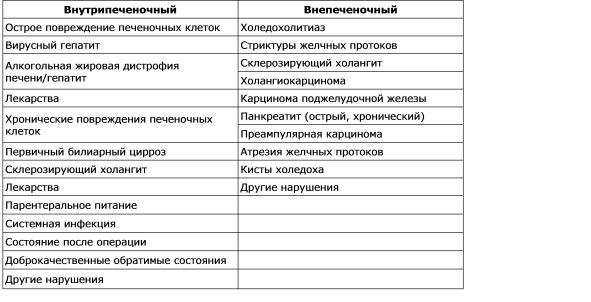
Naruszenie tworzenia się żółci nazywa się cholestazą. Sekwencja pojawiających się patologicznych, fizjologicznych i klinicznych objawów cholestazy zależy od przyczyny. Badanie morfologiczne materiału z żółcią hepatobiopsyjną wykrywa się w kanalikach okołościernych hepatocytów, obserwuje się dylatację kanalików, a badanie ultrastruktury ujawnia zmniejszenie liczby mikrokosmków. Cholestazę można zdefiniować jako defekt funkcjonalny w powstawaniu żółci na poziomie hepatocytów (cholestaza wewnątrzwątrobowa), jak również upośledzenie wydzielania organicznego lub mechanicznego i wypływ żółci (cholestaza pozawątrobowa). Najczęstsze przyczyny cholestazy wewnątrzwątrobowej i pozawątrobowej podano w tabeli. 7-2. Istnieje kilka mechanizmów, które odgrywają ważną rolę w patogenezie cholestazy wewnątrzwątrobowej: upośledzenie i uszkodzenie funkcji błony sinusoidalnej; zakłócenie funkcji wewnątrzkomórkowych organelli hepatocytów; uszkodzenie i rozerwanie membrany rurowej. Tak więc nie ma jednego mechanizmu cholestazy w różnych sytuacjach klinicznych, a wielość mechanizmów może prowadzić do różnych zaburzeń. Klinicznie cholestaza charakteryzuje się wzrostem stężenia wielu substancji we krwi, w tym bilirubiny, soli żółciowych, cholesterolu, które są zwykle wydzielane do żółci. W analizie biochemicznej krwi z zastojem żółciowym obserwuje się nieproporcjonalny wzrost aktywności fosfatazy alkalicznej i stężenia bilirubiny równolegle ze zmianami poziomu aminotransferaz, co zostanie omówione poniżej.
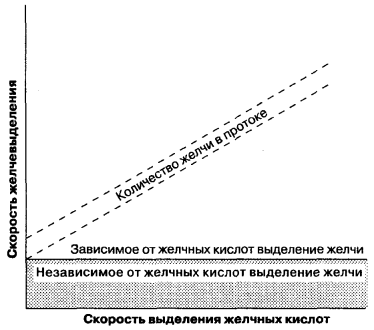
Ryc. 7-5.
Powstanie żółci związane z wydzielaniem kwasu żółciowego, powstawanie żółci niezależne od wydzielania kwasu żółciowego. (Autor: Moseley R. H., wydzielanie żółci, w: Yamada T., Alpers D.H., Owyang C., Powell D. W., Silverstein F.E., wyd. Text of of Gastroenterology, 2. wyd. Philadelphia: J. B. Lippincott, 1995: 387.)
Tabela 7 - 2.
W wątrobie powstaje najważniejszy sok trawienny - żółć.
Żółć jest wytwarzana przez hepatocyty poprzez aktywny i pasywny transport wody, cholesterolu, bilirubiny, kationów do nich. W hepatocytach z cholesterolu powstają pierwotne kwasy żółciowe - choliczne i dezoksycholowe. Kompleks rozpuszczalny w wodzie jest syntetyzowany z bilirubiny i kwasu glukuronowego. Wnikają do kapilar żółciowych i przewodów, w których kwasy żółciowe łączą się z glicyną i tauryną. W wyniku tego powstają kwasy glikocholowe i taurocholowe. Wodorowęglan sodu powstaje w wyniku tych samych mechanizmów, co w trzustce.
Żółć jest wytwarzana przez wątrobę przez cały czas. W jego czasach powstaje około 1 litra. Hepatocyty wydalają pierwotną lub wątrobową żółć. Ciecz ta jest złocistożółtą reakcją alkaliczną. Jego pH = 7,4 - 8,6. Składa się z 97,5% wody i 2,5% substancji stałych. Suchy osad zawiera:
1. Substancje mineralne. Kationy sodu, potasu, wapnia, wodorowęglany, aniony fosforanowe, aniony chloru.
2. Kwasy kwasowe - taurocholowe i glikocholowe.
3. Pigmenty żółciowe - bilirubina i jej utleniona forma biliwerdyna. Bilirubina daje kolor żółci.
4. Cholesterol i kwasy tłuszczowe.
5. Mocznik, kwas moczowy, kreatynina.
Ponieważ poza układem pokarmowym, zwieracz Oddiego, znajdujący się przy ujściu przewodu żółciowego wspólnego, jest zamknięty, wydalana żółć gromadzi się w pęcherzyk żółciowy. Tutaj woda jest z niej ponownie absorbowana, a zawartość podstawowych składników organicznych i mucyny wzrasta 5-10 razy. Dlatego żółć torbielska zawiera 92% wody i 8% suchej pozostałości. Jest ciemniejszy, grubszy i bardziej lepki niż wątroba. Z powodu tego stężenia pęcherz może gromadzić żółć przez 12 godzin. Podczas trawienia dochodzi do otwarcia zwieracza Oddiego i zwieracza z Lutkensa w szyjce pęcherza. Żółć wchodzi do dwunastnicy.
Wartość żółci:
1. Kwasy jabłkowe emulgują część tłuszczów, zamieniając duże cząstki tłuszczu w drobne kropelki.
2. Aktywuje enzymy soku jelitowego i trzustkowego, zwłaszcza lipazy.
3. W połączeniu z kwasami żółciowymi przez błonę enterocytów dochodzi do wchłaniania długołańcuchowych kwasów tłuszczowych i witamin rozpuszczalnych w tłuszczach.
4. Zhelchka promuje resyntezę trójglicerydów w enterocytach.
5. Inaktywuje pepsyny, a także neutralizuje kwaśną miazgę pochodzącą z żołądka. Zapewnia to przejście z trawienia żołądkowego do jelitowego.
6. Pobudza wydzielanie soków trzustkowych i jelitowych, a także proliferację i złuszczanie enterocytów.
7. Wzmacnia ruchliwość jelit.
8. Działa bakteriostatycznie na mikroorganizmy jelitowe i zapobiega powstawaniu w nim procesów gnilnych.
Regulacja wydzielania żółci i wydalanie z żółcią odbywa się głównie za pomocą mechanizmów humoralnych, chociaż nerwowe odgrywają pewną rolę. Najsilniejszym stymulatorem powstawania żółci w wątrobie są kwasy żółciowe, wchłaniane do krwi z jelita. Jest również wzmacniany przez sekretynę, która przyczynia się do wzrostu wodorowęglanu sodu w żółci. Nerw nerwu błędnego stymuluje produkcję żółci, sympatycznego hamowania.
Kiedy treść pokarmowa wchodzi do dwunastnicy, komórki I zaczynają uwalniać swoje komórki cholecystokininy-pankreozyminy. Zwłaszcza ten proces jest stymulowany przez tłuszcze, żółtko jaja i siarczan magnezu. CCK-PZ wzmacnia skurcze mięśni gładkich pęcherza, dróg żółciowych, ale rozluźnia zwieracze z Lutkens i Oddi. Żółć jest uwalniana do jelita. Mechanizmy odruchowe odgrywają niewielką rolę. Chyme podrażnia chemoreceptory jelita cienkiego. Impulsy z nich wchodzą do przewodu pokarmowego rdzenia przedłużonego. Od niego są na vagusie do dróg żółciowych. Zwieracze rozluźniają się, a gładkie mięśnie pęcherza kurczą się. Promuje wydalanie z żółcią.
Najpoważniejsze choroby to zapalenie wątroby i marskość. Najczęściej zapalenie wątroby jest wynikiem zakażenia (zakaźne zapalenie wątroby typu A, B, C) i ekspozycji na produkty toksyczne (alkohol). W zapaleniu wątroby dochodzi do zmian w wątrobie, a wszystkie funkcje wątroby są upośledzone. Marskość jest wynikiem zapalenia wątroby. Najczęstszym naruszeniem wydalania z żółcią jest kamica żółciowa. Większość kamieni żółciowych tworzy cholesterol, ponieważ żółć tych pacjentów jest z nimi przesycona.
Wątroba jest największym narządem w organizmie i ma kluczowe znaczenie dla metabolizmu. Pełni wiele funkcji, uczestnicząc w metabolizmie białek, węglowodanów, tłuszczów, hormonów i witamin, a także w neutralizacji wielu substancji endogennych i egzogennych. Procesy te opisano w podręcznikach chemii fizjologicznej, aw niniejszym rozdziale w związku z układ trawienny rozważymy tylko wydalnicza funkcja wątrobytj. wydzielanie żółci. Żółć składa się z wody, soli mineralnych, śluzu, lipidów cholesterolu i lecytyny oraz dwóch rodzajów specyficznych składników - kwasów żółciowych i pigmentu bilirubiny. Kwasy żółciowe są detergentami, a ich działanie emulgujące odgrywa ważną rolę w trawieniu lipidów. Bilirubina jest końcowym produktem rozkładu hemoglobiny, która ma być usunięta z organizmu.
^
Formacja żółci
Anatomia funkcjonalna.Komórki wątroby (hepatocyty)tworzą płytkę o grubości jednej komórki, rozdzieloną wąskimi szczelinami (Przestrzeń dyskowa)reprezentowanie ukończonych
^ 766 CZĘŚĆ VIII. ŻYWNOŚĆ, TRAWIENIE I EKSTRAKT
Przez krew sinusoidyrównoważny kapilarom. W ścianach sinusoidalnych znajdują się pory, przez które przechodzą tak duże makrocząsteczki, jak albumina i lipoproteiny. W najmniejszym kanalikiżółciowe naczynia krwionośne ograniczone błonami plazmatycznymi dwóch sąsiednich hepatocytów - żółć zbiera się w większej kanał górniczy Goringa,ściany, jak te większe tubulka intercollegei drogi żółcioweutworzone przez sześcienne komórki wydzielnicze. Małe kanaliki wewnątrz płatów wątroby i między nimi łączą się w większe, ostatecznie tworząc przewód wątrobowy.Od tego kanału odchodzi przewód torbielowatydo pęcherzyka żółciowego. Po połączeniu powstają kanały wątrobowe i torbielowate wspólny przewód żółciowyotwarcie w dwunastnicy w górnej części brodawki Vatera, za lub blisko przewodu trzustkowego (ryc. 29.1).
^ Funkcje żółci.Bile wykonuje wiele ważnych funkcji. Z nią wyświetlane są produkty końcowewymiany, np. bilirubiny, a także narkotyków i toksyn. Przydział zżółć cholesterolodgrywa ważną rolę w regulacji jego równowagi. Kwasy żółcioweniezbędny do emulgowania i wchłaniania tłuszczu. Ponadto żółć zawiera wodę, sole mineralne i śluz. Około 24 godziny na dobę 600 mlżółć, a 2/3 tej ilości pochodzi z kanalików i 1/3 większych przewodów.
^ Żółć rurkowa powstaje w przybliżeniu w równych ilościach z udziałem dwóch różnych mechanizmów, zależnych od żółci i zależnych od żółci (ryc. 29.27).
^ Wydzielanie zależne od kwasu żółciowego. Istnieje ścisły związek między szybkość przepływu żółcii wydzielanie kwasu żółciowego.W kanalikowej żółci stężenie kwasów żółciowych jest 100 razy wyższe niż we krwi wrotnej, dlatego uważa się, że są one wydzielane przez aktywny transport z udziałem wektora.Podążając za kwasami żółciowymi wzdłuż gradientu osmotycznego, woda wnika do kanalików, więc żółć jest izotoniczna z krwią.
Istnieją dwa źródła kwasów żółciowych. Po pierwsze, są one syntetyzowane de novo z cholesterolu w samych hepatocytach przy udziale 7-hydroksylazy. Enzym ten odgrywa kluczową rolę i kontroluje szybkość syntezy kwasów żółciowych poprzez mechanizm sprzężenia zwrotnego. Po drugie, hepatocyty są w stanie aktywnie absorbować kwasy żółciowe z krew w portalui wydalają je do kanalików (patrz także rys. 29.29). To ekstrakcja jest bardzo skuteczna; z pojedynczym przepływem krwi przez wątrobę, ekstrahuje się z niego 80% kwasów żółciowych. Dlatego
Stężenie kwasów żółciowych we krwi obwodowej jest znacznie niższe niż w systemie portalowym. Ponieważ kwasy żółciowe są ekstrahowane z krwi 6 razy szybciej niż wchodzą do kanalików, to ten ostatni proces ogranicza szybkość wydzielania kwasu żółciowego.
^ Wydzielanie niezależne od kwasów żółciowych. W
Proces ten obejmuje jony Na +, Cl -, HCO3 - i wodę. Siłą napędową jest transport aktywny Να + , ewentualnie z wodorowęglanem. Wydzielanie niezależne od kwasów żółciowych stymuluje w szczególności sekretyna.
Oprócz kwasów żółciowych bilirubina, cholesterol i fosfolipidy (głównie lecytyna) są aktywnie wydzielane do kanalików (ryc. 29.27). Nierozpuszczalny w wodzie ("Pośrednia") bilirubina,większość z nich jest utworzona z hemoglobiny w wieku czerwonych krwinek, wchodzi do hepatocytów w postaci koloidalnego agregatu związanego z albuminą. Jego codzienne tworzenie wynosi około 4 g / kg masy ciała lub 200-300 mg / dzień. W hepatocytach sprzęga się 80% bilirubiny. zkwas glukuronowy i niewielka jego ilość z kwasem siarkowym. W takim sprzężonym
^ DZIAŁ 29. FUNKCJE UKŁADU GASTRONOMICZNEGO 767
Bilirubina jest wydalana z żółcią. (Bilirubina "bezpośrednia").Ogólnie leki i toksyny są usuwane w ten sam sposób.
^ Modyfikacja żółci w drogach żółciowych
(rys. 29.27). W kanałach, w których kanaliki są otwarte, pierwotna żółć ulega modyfikacji. Proces ten przypomina proces modyfikacji przesączu kłębuszkowego w kanalikach nerkowych (s. 785) iw podobny sposób jest obliczany odprawa,tylko w przypadku żółci jako substancji obojętnej stosuje się zamiast inuliny erytrytollub mannitol,które są wydzielane do kanalików, ale nie są ponownie wchłaniane. Podobne badania wykazały, że około 180 ml żółci lub 1/3 jego całkowitej ilości jest wydzielane do kanałów z aktywną sekrecją NSO ^ "Proces ten jest stymulowany sekretyna
^
Wątroby żółciowej i żółciowej
Skład wątrobowej żółci(tab. 29.3). Żółć wydzielana przez wątrobę z szybkością 0,4 ml / min ma złoty kolor, co tłumaczy się obecnością w niej bilirubiny. Stężenie elektrolitów w tej żółci jest takie samo jak w osoczu, z tym wyjątkiem, że zawiera dwa razy więcej HCO3 "i nieco mniej niż CI ~ .W tym samym czasie, skład substancji organicznych żółci bardzo różni się od osocza, ponieważ w żółci są one reprezentowane prawie wyłącznie przez kwasy żółciowe, cholesterol i fosfolipidy.
 |
^ Kwasy żółciowepowstają w wątrobie z cholesterolu w wyniku jego hydroksylacji i dodania grupy karboksylowej. Powstają kwasy, które tworzą się w wątrobie pierwotne kwasy żółciowe;obejmują one chenodeoksycholowy(dioxyform) i holevaya(trioxyforma) kwas.W wątrobie nie występują one w postaci wolnej, ale w postaci koniugatów z glicyną i tauryną, a koniugaty z glicyną są tworzone trzy razy więcej, ponieważ ilość tauryny jest ograniczona. Sprzężone kwasy żółciowe są bardziej rozpuszczalne w wodzie,
Niż nieskoncentrowane i mają większą zdolność do dysocjacji i tworzenia soli żółciowych z kationami, głównie z jonami Na +. W kwaśnym środowisku (pH 4,0) sole kwasów żółciowych są nierozpuszczalne i wytrącają się, ale przy fizjologicznych wartościach pH (w jelicie cienkim) są one dobrze rozpuszczalne.
W dalszej części jelita krętego i okrężnicy część soli pierwotnych kwasów żółciowych ulega dehydroksylacji pod działaniem bakterii beztlenowych i przekształca się w kwas żółciowy wtórnie-litocholowy(monoetoksyform) i deoksycholowy(dioxyform). Kwasy chinodoksycholowy, cholowy i deoksycholowy są obecne w stosunku 2: 2: 1. Kwas litocholowy występuje tylko w niektórych frakcjach, ponieważ większość z nich jest wydalana.
Emulgujący wpływ kwasów żółciowych na tłuszcze opiera się głównie na ich zdolności do tworzenia micele.Cząsteczki kwasu żółciowego mają trójwymiarową strukturę, w której hydrofilowe grupy karboksylowe i hydroksylowe znajdują się po tej samej stronie cząsteczki, a hydrofobowa część cząsteczki (rdzeń steroidowy, grupy metylowe) znajduje się po przeciwnej stronie, dzięki czemu cząsteczki kwasu żółciowego mają i hydrofilowyi właściwości lipofilowe.Dzięki tej strukturze cząsteczki kwasów żółciowych działają jak detergenty: na granicy fazy lipidowej i wodnej tworzą prawie monomolekularny film, w którym grupy hydrofilowe są przekształcane w fazę wodną, a lipofilową w fazę lipidową. W fazie wodnej kwasy żółciowe tworzą uporządkowane agregaty. -micelspod warunkiem, że ich koncentracja osiągnie określony poziom, zwany krytyczne stężenie miceli(1-2 mmol / l). Wewnętrzny, lipofilowy obszar miceli może zawierać lipidyna przykład cholesterol i fosfolipidy; takie micele są nazywane mieszanymi (ryc. 29.28). Sam cholesterol jest nierozpuszczalny w wodzie, ale może znajdować się w roztworze w składzie miceli. Jeśli jego stężenie przekracza pojemność miceli, tworzy krystaliczny osad; proces ten leży u podstaw powstawania kamieni żółciowych cholesterolu (str. 769).
^ Skład torbielowatej żółci (tab. 29.3). Pojemność pęcherzyka żółciowego wynosi tylko 50-60 ml. W tym samym czasie wątroba wydziela żółć z szybkością 600 ml / dobę, a połowa tej ilości przechodzi przez pęcherzyk żółciowy przed wejściem do jelita cienkiego. Różnica między objętością żółci wchodzącej do pęcherzyka żółciowego a jej wydajnością jest kompensowana wysoce wydajna reabsorpcjawoda w pęcherzyku żółciowym. W ciągu kilku godzin 90% wody może zostać ponownie wchłonięte z żółci. Dzięki
^
768 CZĘŚĆ VIII. ŻYWNOŚĆ, TRAWIENIE I EKSTRAKT
Ta materia organiczna pozostaje w pęcherzyku żółciowym, a ich stężenie w żółci wzrasta. Siłą napędową reabsorpcji jest aktywny transport jonów Na *z udziałem "pompy" wbudowanej w podstawowe i boczne błony komórek i aktywowanej przez (Na + -K +) - ATPazę. Po jonach Na + rozpraszają się jony Cl - i HCO3 - w kierunku gradientu elektrycznego lub transportowane przez ruch nośników. W wyniku reabsorpcji HCO3 - pH pęcherzyka żółciowego zmniejsza się do 6,5 w porównaniu do 8,2 w wątrobie z żółcią. W wyniku wytworzenia się wysokiego stężenia jonów Na + w nabłonku pęcherzyka żółciowego w przestrzeni pozakomórkowej zachodzi gradient osmotyczny, prowadzący do pompowania wody, która następnie wpływa do naczyń włosowatych (s. 751).
^ Ruchliwość pęcherzyka żółciowego. W stanie na czczo żółć gromadzi się w woreczku żółciowym i w
Czas jedzenia jest przydzielany w wyniku skurczów pęcherzyka żółciowego. Głównym stymulatorem kurczliwości pęcherzyka żółciowego jest cholecystokinina,wydzielany przez błonę śluzową dwunastnicy przy przyjęciu do niej mięty zawierającej tłuszcz. Skurcze pęcherzyka żółciowego są również do pewnego stopnia stymulowane. błąkający się nerwi parasympatolityki.Zaczynają się w ciągu 2 minut po zetknięciu się tłuszczu z błoną śluzową jelita, a po 15-90 minutach pęcherz zostaje całkowicie opróżniony. Ruchliwość pęcherzyka żółciowego obejmuje dwa procesy. Początkowo rozwija się skurcz toniczny, w wyniku czego zmniejsza się średnica pęcherzyka żółciowego, a następnie na ten efekt nakładane są okresowe skurcze, których częstotliwość wynosi 2-6 / min. W wyniku tych dwóch procesów powstaje ciśnienie 25-30 mm Hg. Art.
^
Krążenie wątrobowo-wątrobowe
Krążenie kwasów żółciowych(rys. 29.29). Kwasy żółciowe są wydzielane do dwunastnicy w postaci mieszanych miceli. Pomimo rozcieńczenia kwasów żółciowych przez zawartość żołądka, ich stężenie w jelicie wynosi około 10 mmol / l i pozostaje powyżej krytycznego stężenia tworzenia się miceli. Tutaj, obok cholesterolu i lecytyny, micele obejmują produkty hydrolitycznego rozpadu tłuszczu - kwasy tłuszczowei monoglicerydy.Przy początkowym kontakcie miceli ze ścianą jelita
^
DZIAŁ 29. FUNKCJE UKŁADU GASTRONOMICZNEGO 769
Lipidy dyfundują przez błonę granicy szczoteczki do enterocytów, a kwasy żółciowe pozostają w świetle jelita, ale przy dalszym przejściu przez jelita kwasy żółciowe są absorbowane przez aktywny i pasywny transport.
Około 50% kwasów żółciowych jest pasywnie absorbowanych w jelicie. W wyniku rozszczepienia koniugatów kwasów żółciowych i dehydroksylacji tych ostatnich przez działanie bakterii jelitowych, zwiększa się ich rozpuszczalność w lipidach i ułatwia się dyfuzję bierną.
^ Aktywne ssanie kwasy żółciowe występują wyłącznie w końcowym odcinku jelita krętego - rzadkim zjawisku, znanym jedynie z wchłaniania witaminy B12. Tylko te kwasy żółciowe, które mają wysoką polarność, która utrudnia ich bierne wchłanianie, takie jak koniugaty tauryny, podlegają aktywnej absorpcji. Proces wchłaniania kwasów żółciowych w końcowym odcinku jelita krętego charakteryzuje się typowymi oznakami aktywnego transportu: kinetyką nasycenia i konkurencyjnym hamowaniem. Niewielka ilość kwasów żółciowych (7-20%) nie jest włączona w absorpcję czynną ani bierną i jest eliminowana z organizmu.
Obecność kwasów żółciowych w okrężnicy wydaje się odgrywać ważną rolę w regulowaniu konsystencji kału. Gdy stężenie Dioxy Acid w okrężnicy przekracza 3 mmol / l, znaczna ilość elektrolitów i wody jest wydzielana do światła jelita, co prowadzi do biegunki. Wyraźna forma tego Biegunka "Hologennogo"można obserwować po resekcji lub chorobie kończyn dolnych jelit, a także w leczeniu za pomocą wiązania kwasów żółciowych za pomocą wymiennika jonowego cholestyraminy.
Po spożyciu wchłania kwasy żółciowe do wątrobykoniugaty są nowo powstałe, a niektóre drugorzędowe kwasy żółciowe ulegają hydroksylacji. Utrata kwasów żółciowych w kale (0,2-0,6 g / dzień) jest kompensowana przez ich syntezę.
^ Całkowita pula kwasów żółciowych w organizmie wynosi około 3,0 g. Ta ilość nie jest wystarczająca do zapewnienia lipolizy po posiłku; w szczególności spożywanie tłustych potraw wymaga 5 razy więcej kwasów żółciowych. Jednak organizm nie wykazuje niedoboru kwasów żółciowych, ponieważ krążą one wielokrotnie w jelitach i wątrobie. (krążenie jelitowo-wątrobowe).Częstotliwość, z jaką pula kwasów żółciowych tworzy pełny cykl, zależy od reżimu dietetycznego i waha się od 4 do 12 cykli dziennie.
^ Krążenie bilirubiny. Pigment galusowy bilirubina,podobnie jak kwasy żółciowe i lipidy, dostaje się do jelita w postaci glwuronidu. Tylko niewielka ilość tego związku polarnego
Reabsorbowane w pęcherzyku żółciowym i jelicie cienkim. W końcowym odcinku jelita krętego i (głównie) w okrężnicy koniugat bilirubiny jest odcinany przez działanie hydrolaz bakteryjnych. W tym samym czasie zamienia się bilirubina urobilinogenktóry, wraz z innymi produktami rozpadu bilirubiny, nadaje brązowej barwie kał. Mniej niż 20% urobilinogenu jest absorbowane z powrotem, a tej ilości około 90% ponownie wchodzi do wątroby i wraca do żółci, a pozostałe 10% jest wydalane z moczem.
Aspekty patofizjologiczne. Może to wskazywać na podwyższony poziom urobilinogenu w moczu choroba wątroby,towarzyszy temu naruszenie wydalania bilirubiny. Całkowity brak urobilinogenu w moczu, jasny kolor kału i zażółcenie wskazują całkowita blokada pęcherzyka żółciowego;w tej sytuacji bilirubina w ogóle nie dostaje się do jelita, a urobilinogen się nie tworzy.
Najbardziej znanym i szeroko rozpowszechnionym naruszeniem normalnej fizjologii układu żółciowego jest wytrącanie cholesterolu z utworzeniem cholesterolowych kamieni żółciowych. Cholesterol, podobnie jak lecytyna, jest w stanie rozpuszczonym tylko jako część mieszanych miceli. Jeśli stężenie wzrośnie xo.jecmepo.ia albo stężenie kwasów żółciowych lub lecytyny spada poniżej poziomów krytycznych,wydzielanie cholesterolu. Wśród czynników powodujących wzrost względnej zawartości cholesterolu są estrogeny, dieta węglowodanowa, nadwaga i procesy zmniejszające stężenie kwasów żółciowych, takie jak zapalenie jelita krętego. (Choroba Leśniowskiego-Crohna)lub resekcja. W niektórych przypadkach doustna suplementacja kwasem żółciowym może wystarczyć, aby litogeniczna żółć zamieniła się w alitogenną, w której kamienie cholesterolu mogą się rozpuszczać. Do tego celu najlepiej nadają się kwasy: Chenodeoksycholowy i urodesoksycholowy, ponieważ nie powodują biegunki.
Klinicznym objawem upośledzonego metabolizmu bilirubiny jest żółtaczka. Żółta skóra jest związana ze zwiększonym stężeniem bilirubiny w osoczu, co może wystąpić w następujących przypadkach:
ze zwiększonym tworzeniem się bilirubiny w wyniku zwiększonego rozpadu erytrocytów (żółtaczka hemolityczna);
w wyniku przerwania procesu koniugacji lub transportu bilirubiny w hepatocytach, jak na przykład żółtaczka w ciążylub Żółtaczka Gilberta;
z opóźnionym odpływem żółci, na przykład z powodu kamieni żółciowych lub nowotworów zlokalizowanych w rejonie przewodu żółciowego (żółtaczka zaporowa).