La formazione della bile. La fisiologia della formazione della bile. Formazione di bile
La formazione della bile si verifica nel fegato a causa del trasporto attivo e passivo di sostanze (acqua, glucosio, creatinina, elettroliti, ormoni, vitamine, ecc.) Che circolano nel sangue, attraverso le cellule e i contatti cellulari, così come la secrezione attiva dei componenti biliari (acidi biliari) da epatociti e riassorbimento di acqua e alcune sostanze dai piccoli dotti biliari e calcoli biliari. Il significato fisiologico di questo processo è vario. La bile è principalmente considerata un segreto digestivo, poiché gli acidi biliari (principalmente i loro anioni organici) svolgono un ruolo chiave nell'assorbimento dei grassi. La bile emulsiona i grassi, aumentando la superficie su cui vengono idrolizzati sotto l'effetto della lipasi. Sotto l'azione della bile c'è la dissoluzione dei prodotti grassi di idrolisi. Promuove il loro assorbimento e risintesi dei trigliceridi negli enterociti. La bile aumenta l'attività degli enzimi del pancreas e dell'intestino (specialmente le lipasi), migliora l'idrolisi e l'assorbimento di proteine e carboidrati.
In violazione della digestione dei grassi sono mal digeriti e altre sostanze alimentari, perché il grasso avvolge le più piccole particelle di cibo e impedisce l'azione degli enzimi su di loro. In tali condizioni, l'attività dei batteri intestinali porta ad un aumento dei processi di decadimento, fermentazione e formazione di gas.
Effetti intrinseci e regolatori della bile: stimolazione della formazione di bile, escrezione biliare, attività motoria e secretoria dell'intestino tenue, nonché proliferazione e desquamazione degli enterociti. La bile inibisce il processo di digestione gastrica neutralizzando l'acido che è entrato nel duodeno e inattivando la pepsina, preparando la digestione a livello intestinale. Ha anche un impatto normativo sull'attività di evacuazione del complesso gastroduodenale. Il ruolo della bile nell'assorbimento delle vitamine liposolubili (A, D, E e K), il colesterolo, i sali di calcio dal lume intestinale è importante.
Inoltre, la formazione e la secrezione della bile sono considerate un modo peculiare di escrezione di alcune molecole e ioni che non possono essere espulsi attraverso i reni. Tra questi, i più significativi sono il colesterolo (derivato dal colesterolo libero, i suoi esteri e acidi biliari), la bilirubina, nonché le molecole di rame e ferro. Pertanto, la bile è considerata come un fluido escretore.
La bile consiste di 80% di acqua e 20% di sostanze disciolte in esso. Questi ultimi includono acidi biliari e loro sali (circa il 65%), fosfolipidi biliari (circa il 20%, principalmente a causa della lecitina), proteine (circa il 5%), colesterolo (4%), bilirubina coniugata (0,3%), enzimi , immunoglobuline, nonché numerose sostanze esogene ed endogene secrete dalla bile (stirene vegetale, vitamine, ormoni, farmaci, tossine, ioni metallici - rame, ferro, potassio, sodio, calcio, zinco, magnesio, mercurio, ecc.). In media vengono secreti 600-700 ml di bile (da 250 a 1500 ml, circa 10,5 ml per 1 kg di peso corporeo). Allo stesso tempo, circa 500 ml / die del volume totale sono forniti dalla secrezione di epatociti e circa 150 ml / die le cellule dei dotti biliari secernono.
Formazione di bile (Cholepoiesis) va continuamente, e il flusso della bile nel duodeno (Holekinez) succede periodicamente. A stomaco vuoto, la bile entra nell'intestino secondo un'attività periodica affamata. Durante i periodi di riposo, va al WB, dove si concentra, cambia leggermente la sua composizione e viene depositato. Oltre all'acqua e ai sali, il colesterolo e gli acidi grassi liberi vengono assorbiti. A questo proposito, distinguere la bile epatica e della cistifellea.
La bile ha una piccola attività enzimatica; Il pH della bile epatico è 7.3-8.0. A differenza dei contenuti intestinali, contiene quasi nessun batterio. I fattori che assicurano la sterilità della bile comprendono la presenza di acidi biliari (effetto batteriostatico), il contenuto ricco di immunoglobuline, la secrezione di muco, la relativa povertà dei substrati di energia bile per i batteri.
La bile è una soluzione micellare. Il colesterolo, praticamente insolubile in acqua, viene trasportato in uno stato disciolto nella bile a causa della sua struttura micellare. Questo processo è chiamato dissoluzione colloidale - solubilizzazione.
Gli acidi biliari sono molecole tensioattive, anfipatiche (sia idrofobe che idrofile) in grado di autoaggregarsi. In questo caso, grazie a un tasso di concentrazione molto stretto, chiamato concentrazione critica di micellizzazione, si formano semplici micelle. Le micelle semplici hanno una spiccata capacità di dissolvere i lipidi, formando micelle miste.
Si ritiene che le micelle miste abbiano una struttura cilindrica: il gambo cilindrico è riempito di lipidi polari e le molecole di acido biliare si trovano tra le estremità polari di molecole lipidiche con lati idrofili rivolti verso l'ambiente acquatico, che ne determinano l'idrofilia (solubilità in acqua).
Le micelle miste contengono componenti essenziali - acidi biliari, situati all'esterno, fosfolipidi (principalmente fosfatidilcolina - lecitina) e colesterolo, situati all'interno della micella.
Il colore della bile è di colore bruno-giallastro a causa della presenza di bilirubina, la maggior parte delle quali viene presentata sotto forma di bilirubina diglucuronide, la più piccola - sotto forma di bilirubina monoglucuronide. La bilirubina associata non fa parte delle micelle.
Poiché l'albero biliare intercellulare è permeabile all'acqua, la cistifellea e la bile epatica sono isotoniche.
Formazione di bile
Secrezione biliare
Gli acidi biliari sono il componente principale della secrezione biliare, si formano solo nel fegato. Acidi biliari primari - triidrossicolico (cholio) e diidrossicolico (chenodeossicolico) sono sintetizzati negli epatociti di colesterolo. Acidi biliari secondari (desossicolico e in piccole quantità - litocolico) si formano nel colon dal primario a seguito di modificazione batterica di gruppi idrossilici nucleari (7a-deidrossilazione sotto il controllo di batteri anaerobici). Acidi biliari terziari (principalmente ursodesossicolico) si formano nel fegato per isomerizzazione degli acidi biliari secondari.
La sintesi degli acidi biliari dal colesterolo è regolata da un meccanismo di feedback negativo: un ritorno ridotto degli acidi biliari agli epatociti con sangue portale porta ad un aumento della loro biosintesi; qualsiasi aumento della sintesi degli acidi biliari è accompagnato da un corrispondente aumento della formazione di colesterolo.
Gli acidi biliari sono coniugati nel fegato con gli amminoacidi glicina (circa l'80%) o la taurina (circa il 20%). Quando si consumano cibi ricchi di carboidrati, la quantità di acidi glicocolic aumenta nella bile e nelle diete ad alto contenuto proteico, quelle taurocoliche. Il legame all'amminoacido impedisce l'assorbimento degli acidi biliari nelle vie biliari e nelle sezioni iniziali dell'intestino tenue (solo nell'ileo terminale e nell'intestino crasso). Sotto l'azione dei batteri, è possibile l'idrolisi dei sali biliari con la formazione di acidi biliari, glicina o taurina.
Dopo la biosintesi degli acidi biliari, come notato sopra, il loro gruppo carbossilico è legato al gruppo amminico di glicina o taurina. Da un punto di vista chimico, tale legame converte un acido debole in uno più forte. Gli acidi biliari legati sono più solubili a pH basso e più resistenti alle precipitazioni con gli ioni di calcio (Ca 2+) rispetto agli acidi biliari non legati. Dal punto di vista biologico, il processo di legame rende gli acidi biliari incapaci di penetrazione attraverso le membrane cellulari. Pertanto, essi vengono assorbiti nei dotti biliari o nell'intestino tenue, sia in presenza di molecole carrier, sia se gli acidi biliari subiscono una scissione batterica. L'unica eccezione a questa regola sono gli acidi biliari associati alla glicina diidrossi, poiché possono essere assorbiti passivamente se ricevono uno ione idrogeno (H +).
La maggior parte degli acidi biliari (circa 85-90%), ricevuti con il flusso della bile nell'intestino tenue, vengono assorbiti nel sangue. Tuttavia, la maggior parte degli acidi biliari associati che sono secreti nell'intestino tenue vengono assorbiti intatti. Una parte più piccola dell'acido non viene assorbita intatta, poiché subisce una scissione batterica nell'intestino tenue distale. Sono assorbiti passivamente e, ritornando al fegato, sono nuovamente legati e secreti nella bile. Il restante 10-15% degli acidi biliari viene eliminato dal corpo principalmente con le feci. Questa perdita di acidi biliari è compensata dalla loro sintesi in epatociti.
Questo processo di scissione intestinale e riassorbimento nel fegato è una parte normale del metabolismo degli acidi biliari. Una piccola porzione di acidi biliari, non assorbita, entra nel colon. Qui finisce il processo di divisione. Inoltre, gli acidi biliari secondari si formano nel colon sotto l'azione dei batteri anaerobici (vedi sopra).
Gli acidi deoxycholic e lithocholic sono parzialmente assorbiti nel colon e rientrano nel fegato. Dopo il ritorno, il metabolismo di questi acidi biliari è diverso. L'acido desossicolico si lega alla glicina o alla taurina e circola con gli acidi biliari primari. Va notato che negli anziani, l'acido desossicolico è l'acido biliare principale nella composizione della bile. L'acido litocholico si lega non solo alla glicina o alla taurina, ma in aggiunta ai solfati nella posizione C-3. Tale legame "doppio" riduce la possibilità di assorbimento nell'intestino, a causa della quale l'acido litocolico viene rapidamente perso dalla composizione degli acidi biliari circolanti - il suo contenuto nella bile raramente supera il 5%. La maggior parte degli acidi biliari (95%), che fanno parte della bile degli adulti, sono acidi colico, chenodesossicolico e desossicolico.
Il processo di legame degli acidi biliari negli epatociti è molto efficace, quindi gli acidi biliari biliare sono completamente presenti in forma coniugata. La successiva scissione batterica e la deidrossilazione fanno sì che gli acidi biliari nelle feci siano nello stato non legato.
Il processo di deidrossilazione degli acidi biliari primari sopra descritto riduce la loro capacità di dissolversi in acqua. La formazione di acidi biliari secondari nell'uomo non ha praticamente alcun significato fisiologico, a differenza degli animali. Eccessivo assorbimento di acido desossicolico nel colon aumenta il rischio di calcoli biliari di colesterolo. L'acido litocholico è considerato epatotossico. Negli esperimenti sugli animali, è stato dimostrato che l'accumulo di acido litocolico nel fegato porta alla sua sconfitta. Tuttavia, non è stato ancora dimostrato che un aumento nell'assorbimento di questo acido nell'uomo porti all'interruzione del fegato.
L'acido ursodesossicolico e l'acido desossicolico si lega al fegato e circola con gli acidi biliari primari. Tuttavia, la via metabolica di questo acido è molto breve e il contenuto di acido ursodesossicolico nella bile non supera mai il 5% della quantità totale di acidi biliari. Si ritiene che la formazione di acido ursodesossicolico non abbia un importante significato fisiologico.
Gli acidi biliari sono potenti solventi dei grassi, pertanto sono citotossici in concentrazioni che raggiungono la concentrazione critica di micellizzazione. Allo stesso tempo, gli acidi chenodesossicolici e desossicololici presenti nella bile sono più tossici di quelli associati a colica e ursodesossicolico. Sebbene gli acidi biliari siano in modo dimostrativo citotossico in vitro, l'epitelio dei dotti biliari e dell'intestino tenue non viene mai danneggiato a causa della loro elevata concentrazione. Questo è associato, in primo luogo, alla presenza di altri lipidi (fosfolipidi nella bile e acidi grassi nell'intestino), che riducono la concentrazione monometrica degli acidi biliari e, in secondo luogo, con la presenza di glicolipidi e colesterolo nelle membrane apicali delle cellule epiteliali, che aumentano la resistenza delle cellule epiteliali al danneggiamento gli effetti degli anioni degli acidi biliari.
Gli acidi colico, chenodesossicolico e desossicolico vengono assorbiti e sottoposti a circolazione enteroepatica fino a 6-10 volte al giorno. L'acido litocholico è scarsamente assorbito e la sua quantità nella bile è piccola. Il pool di acidi biliari è normalmente di circa 2,5 g, e la produzione giornaliera di acidi biliari primari, cholica e chenodeossicolico, è in media di circa 330 e 280 mg, rispettivamente.
Regolazione della formazione della bile
Come è noto, la formazione della bile avviene continuamente, ma l'intensità di questo processo varia. Migliora l'assunzione di cibo nella formazione della bile e cibo accettato. L'effetto riflesso sulla coleresi si verifica durante la stimolazione dei recettori del tratto gastrointestinale (GIT), degli organi interni e degli effetti riflessi condizionati.
La regolazione vegetativa è fornita da colinergici parasimpatici (aumento della formazione di bile) e fibre nervose simpatiche adrenergiche (riduzione della formazione di bile).
La regolazione umorale è effettuata dalla stessa bile a causa della presenza di circolazione enteroepatica di acidi biliari e del meccanismo di feedback negativo (vedi sopra). La secretina stimola la secrezione della bile, la secrezione di acqua ed elettroliti nella sua composizione. Il glucagone, la gastrina e la colecistochinina hanno un effetto stimolante più debole.
Escrezione biliare
L'escrezione biliare è considerata un modo particolare per rimuovere le molecole e gli ioni dal corpo che non possono essere espulsi attraverso i reni. I più importanti tra questi sono il colesterolo (come tale e sotto forma di acidi biliari) e la bilirubina, così come gli ioni di rame, ferro, ecc.
I componenti principali dell'escrezione biliare
colesterolo, praticamente insolubile in acqua, viene trasportato nella composizione di micelle miste formate, come sopra notato, da acidi biliari, fosfolipidi e colesterolo stesso.
Le molecole fosfolipidi svolgono due importanti funzioni. Innanzitutto, aumentano significativamente la solubilità micellare del colesterolo, poiché le micelle miste contenenti fosfolipidi solubilizzano molto più colesterolo delle micelle semplici contenenti solo molecole di acido biliare. In secondo luogo, la presenza di fosfolipidi nella bile riduce la concentrazione critica di micellizzazione e la concentrazione monometrica degli acidi biliari. Di conseguenza, l'attività superficiale e la citotossicità della bile epatica sono ridotte.
Con il flusso della bile, le micelle miste entrano nell'intestino tenue, dove l'ulteriore trasformazione delle loro componenti costitutive è diversa. Gli acidi biliari sciolgono i lipidi, assicurando il loro assorbimento e vengono assorbiti in più parti distali dell'intestino. Fosfolipidi biliari, insolubili in acqua, idrolizzano nell'intestino e non sono coinvolti nella circolazione enteroepatica. Gli acidi biliari regolano la loro escrezione e stimolano la sintesi.
Poiché circa 1/3 del colesterolo viene assorbito, 2/3 di esso vengono escreti. In un adulto, l'equilibrio del colesterolo è assicurato dal suo rilascio in quanto tale (circa 600 mg / giorno) o come acidi biliari (circa 400 mg / giorno). Allo stesso tempo, la circolazione enteroepatica degli acidi biliari è considerata una via ritardata di escrezione di colesterolo.
Va notato che, rispetto agli animali, l'uomo ha una percentuale maggiore di colesterolo nella bile. Questo è considerato il risultato di una conversione difettosa del colesterolo agli acidi biliari, così come il tasso relativamente basso di secrezione degli acidi biliari. Pertanto, la bile è sovrasaturi di colesterolo in circa il 25% delle persone anziane e nel 10-15% delle pietre di colesterolo della popolazione (colelitiasi) si formano.
Come notato sopra, la sintesi degli acidi biliari dal colesterolo è regolata da un meccanismo di feedback negativo.
La bile è il modo principale per rimuovere la bilirubina dal corpo. bilirubina - Il prodotto finale della decomposizione dell'eme, nella sua struttura chimica è il tetrapirrolo. La più grande quantità di bilirubina (80-85%) proviene dalla decomposizione dell'emoglobina dei globuli rossi invecchiati e dal prematuro collasso dei globuli rossi appena formati nel midollo osseo o nel letto circolatorio (la cosiddetta eritropoiesi inefficace). Il resto della bilirubina si forma come risultato della distruzione di altre proteine contenenti ormoni (ad esempio, citocromo P-450, ecc.) Nel fegato e molto meno nel tessuto extraepatico. Va notato che il catabolismo dell'emoglobina eritrocitaria in bilirubina si verifica principalmente nei macrofagi della milza, del fegato e del midollo osseo.
Fonti di formazione di bilirubina (in percentuale)

Come è noto, la bilirubina non coniugata è idrofobica (insolubile in acqua) e una sostanza potenzialmente tossica che circola nel plasma in uno stato strettamente correlato all'albumina e non può essere escreta nelle urine. La capacità del corpo di rimuovere la bilirubina è associata alla rimozione di quest'ultimo dal plasma sanguigno dalla cellula epatica, successiva coniugazione con acido glucuronico e rilascio nella bile di un composto già idrosolubile (bilirubina legata o diretta). Il processo di coniugazione procede attraverso l'enzima microsomiale uridina difosfato glucuronil transferasi. La coniugazione della bilirubina con l'acido glucuronico ha il più importante significato fisiologico, tuttavia la coniugazione della bilirubina con solfati, glucosio e xilosio avviene in piccole quantità.
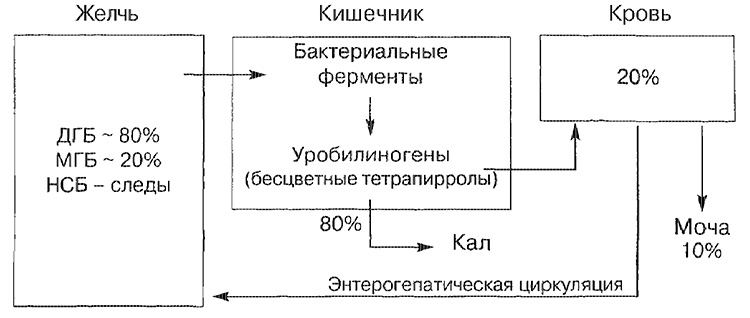
Principali vie di trasporto per la bilirubina
NSB - bilirubina non legata; MGB - bilirubina monoglucuronide; DGB - bilirubina diglucuronide.
La secrezione di bilirubina coniugata nei tubuli della bile avviene con la partecipazione di una famiglia di proteine di trasporto multispecifico ATP-dipendenti per anioni organici.
La maggior parte della bilirubina bilirubina (circa 80%) è rappresentata nella forma di bilirubina diglucuronid, una parte più piccola - nella forma di un monoglucuronide, e solo una piccola parte di essa è rappresentata da una forma non legata. La bilirubina, che è entrata nell'intestino, è scomposta nella parte terminale dell'intestino tenue e nell'intestino crasso da enzimi batterici (beta-glucuronidasi), che viene convertita in tetrapirrolo incolore (urobilinogeno). Circa il 20% degli urobilinogeni formati vengono riassorbiti e necessariamente escreti nelle urine, così come nella bile (circolo enteroepatico di circolazione).
La maggior parte degli autori concorda che la normale concentrazione di bilirubina sierica totale, determinata dalla reazione diazoica di Van Der Bergh, di solito non supera 1 mg% (0,3-1 mg% o 5-17 μmol / l). Solo meno del 5% della bilirubina è presentato in forma legata. Un aumento del livello di bilirubina nel sangue (iperbilirubinemia) e il suo accumulo nei tessuti portano alla comparsa di ittero, che, di regola, diventa evidente a valori superiori a 2,5-3 mg%.
Come notato sopra, la bilirubina circola nel sangue in uno stato legato alle proteine e penetra a fatica nei fluidi tissutali con un basso contenuto proteico. A questo proposito, gli essudati sono di colore più giallo rispetto ai trasudati. La bilirubina si lega bene ai tessuti elastici, il che spiega la precoce colorazione gialla della sclera, della pelle e delle pareti vascolari nell'iperbilirubinemia. La colestasi prolungata porta alla comparsa di un tono della pelle verdastro, che è spiegato dalla deposizione di biliverdina.
Sarà opportuno sottolineare che la pressione nei dotti biliari, sullo sfondo di cui si verifica la secrezione biliare, normalmente corrisponde a 150-200 mm di acqua. Art. Il suo aumento a 350 mm di acqua. Art. inibisce la secrezione della bile, portando allo sviluppo di ittero. Nel caso di completa cessazione della secrezione di bilirubina e acidi biliari, la bile diventa scolorita (la cosiddetta bile bianca).
Inoltre, la bile è un modo di escrezione grassi vegetali, farmaci lipofili e loro metaboliti, vari xenobiotici presenti nelle piante, metaboliti lipofili, vitamine liposolubili e ormoni steroidei.
Equilibrio di ferro e rame il corpo è anche supportato a causa dell'escrezione biliare di questi metalli. Entrambi i cationi sono secreti nella bile da una pompa canalicolare stimolata da ATP. Con la bile si verifica e il rilascio di altri metalli.
Escrezione di bile
La corrente della bile nel sistema biliare è causata dalla differenza di pressione nelle sue varie sezioni e duodeno, tono dello sfintere, contrazione delle fibre muscolari lisce del wormhole e dei dotti (le prime fibre muscolari individuali compaiono nei dotti biliari intraepatici situati nelle immediate vicinanze delle porte del fegato). L'attività dell'intero sistema è normalmente ben coordinata e sotto il controllo dei meccanismi nervoso e umorale.
Nei calcoli biliari, le concentrazioni di bile si depositano e aumentano con un aumento di 10 volte della concentrazione di lipidi, sali biliari, pigmenti biliari, ecc. Al momento giusto, viene rilasciato nell'OZhP e più avanti nel duodeno. Il passaggio della bile coordina l'apparato sfinteriale. Al di fuori del processo digestivo nel duodeno, lo sfintere della fiala viene chiuso. A questo punto, il RI e il dotto cistico sono rilassati, il che consente alla bile prodotta dal fegato di entrare nel lume del RI. La membrana mucosa ZH assorbe acqua, ioni. Allo stesso tempo la bile diventa più concentrata. I prodotti del muco permettono alla bile di essere in uno stato colloidale.
La pressione nel lume della ghiandola a riposo è molto inferiore rispetto ai dotti biliari, ed è di 60-185 mm di acqua. Art. La differenza di pressione è la base fisiologica del flusso di bile nella cistifellea con lo sfintere Oddi chiuso. Nel processo di digestione a causa della riduzione della pressione GF aumenta a 150-260 mm di acqua. st, assicurando il flusso della bile nel duodeno attraverso le fiale rilassate dello sfintere. Quando la bile inizia a fluire nel duodeno, la pressione nei condotti diminuisce gradualmente (ad ogni pasto, il contenuto di grasso diminuisce 1-2 volte).
Il periodo della reazione primaria del sistema biliare, a causa del tipo, odore del cibo e la sua ricezione, dura circa 7-10 minuti. Poi arriva il periodo di evacuazione (il principale o il periodo di svuotamento dello ZHP), durante il quale, sullo sfondo dell'alternanza di contrazioni e rilassamenti della ZP, la cistifellea e poi la bile epatica sfociano nel duodeno.
La stimolazione dei riflessi di Cholekinesis (riflesso condizionato e incondizionato) avviene a causa di recettori localizzati nella cavità orale, nello stomaco e nel duodeno, attraverso i nervi vago. La regolazione umorale viene effettuata principalmente sotto l'influenza della colecistochinina, che ha un effetto stimolante sul GF, causandone la riduzione. Inoltre, i tagli deboli in GI si verificano sotto l'influenza di gastrina, secretina e Bombezin. Al contrario, glucagone, calcitonina, anticholecistochinina, peptide vasointestinale e polipeptide pancreatico inibiscono la riduzione del GI.
I tuorli d'uovo, i grassi, il latte e la carne sono potenti stimolanti dell'escrezione biliare.
Disturbi dell'apparato sfinterico e motilità gonadica possono causare cambiamenti nella direzione e velocità del flusso biliare, discinesia, reflusso delle secrezioni pancreatiche e il contenuto di duodeno negli ACP, e la bile nel dotto pancreatico, che è la base per lo sviluppo di una serie di stati patologici e malattie.
Sali biliari, bilirubina coniugata, colesterolo, fosfolipidi, proteine, elettroliti e acqua sono secreti dagli epatociti nei canalicoli biliari. L'apparato di secrezione biliare include proteine di trasporto membrana tubolare, organelli intracellularie strutture citoscheletro. Contatti strettitra gli epatociti, il lume dei tubuli è separato dal sistema circolatorio del fegato.
La membrana tubolare contiene proteine di trasporto per acidi biliari, bilirubina, cationi e anioni. Microvilli aumenta la sua area. Gli organelli sono rappresentati dall'apparato di Golgi e dai lisosomi. Con l'aiuto di vescicole, le proteine (es. IgA) vengono trasportate dalla membrana sinusoidale alla membrana canalicolare, il trasporto delle proteine di trasporto sintetizzate nella cellula per il colesterolo, i fosfolipidi e, possibilmente, gli acidi biliari dai microsomi alla membrana tubulare.
Il citoplasma dell'epatocita attorno al tubulo contiene la struttura del citoscheletro: microtubuli, microfilamentie filamenti intermedi.
I microtubuli sono formati dalla polimerizzazione della tubulina e formano una rete all'interno della cellula, specialmente vicino alla membrana basolaterale e all'apparato di Golgi, prendendo parte al trasporto vescicolare mediato dal recettore, alla secrezione di lipidi e, in determinate condizioni, agli acidi biliari. La formazione di microtubuli è inibita dalla colchicina.
Interacting polimerizzato (F) e actina libera (G) partecipano alla costruzione di microfilamenti. I microfilamenti, concentrandosi intorno alla membrana tubolare, determinano la contrattilità e la motilità dei tubuli. La falloidina, che potenzia la polimerizzazione dell'actina e la citocalasina B, che la indebolisce, inibisce la motilità dei tubuli e causa la colestasi.
I filamenti intermedi sono costituiti da citocheratina e formano una rete tra le membrane plasmatiche, il nucleo, gli organelli intracellulari e altre strutture del citoscheletro. La rottura dei filamenti intermedi porta alla rottura dei processi di trasporto intracellulare e alla obliterazione del lume dei tubuli.
L'acqua e gli elettroliti influenzano la composizione della secrezione tubulare, penetrando attraverso stretti contatti tra gli epatociti a causa del gradiente osmotico tra il lume tubulare e gli spazi di Disse. (corrente paracellulare).L'integrità dei contatti stretti dipende dalla presenza sulla superficie interna della membrana plasmatica della proteina ZO-1 con una massa molecolare di 225 kDa. La rottura di contatti stretti è accompagnata dall'ingresso di molecole più grandi disciolte nei canalicoli, che porta alla perdita del gradiente osmotico e allo sviluppo di colestasi. Allo stesso tempo si può osservare rigurgito della bile canalicolare nella sinusoide.
I canalicoli biliari fluiscono nei dotti, talvolta chiamati colangioli o canali di Goering. Le duttule si trovano principalmente nelle aree del portale e fluiscono nei dotti biliari interlobulari, che sono i primi dei tratti biliari accompagnati da rami dell'arteria epatica e della vena porta e si trovano nella composizione delle triadi portale. I dotti interlobulari, che si fondono, formano i dotti settali fino a che due principali condotti epatici emergono, emergendo dai lobi destra e sinistra nella fessura del portale del fegato.
Secrezione biliare
La formazione della bile avviene con la partecipazione di numerosi processi di trasporto volatili. La sua secrezione è relativamente indipendente dalla pressione di perfusione. La corrente totale della bile nell'uomo è di circa 600 ml / giorno. Gli epatociti forniscono la secrezione di due frazioni di bile: dipendenti da acidi biliari ("225 ml / die) e non dipendenti da questi (" 225 ml / die). I restanti 150 ml / giorno sono secreti dalle cellule del dotto biliare.
La secrezione di sali di acidi biliari è il fattore più importante nella formazione della bile (frazione dipendente dagli acidi biliari).L'acqua si muove sulla scia di sali osmoticamente attivi di acidi biliari. I cambiamenti nell'attività osmotica possono regolare il flusso di acqua nella bile. Esiste una chiara correlazione tra la secrezione di sali degli acidi biliari e il flusso della bile.
L'esistenza della frazione bile, che non dipende dagli acidi biliari, è dimostrata dalla possibilità della formazione di bile che non contiene sali di acidi biliari. Quindi, è possibile continuare il flusso della bile, nonostante l'assenza di escrezione di sali di acidi biliari; la secrezione di acqua in questo caso è dovuta ad altre sostanze solubili osmoticamente attive, come glutatione e bicarbonati.
Meccanismi cellulari della secrezione biliare
Un epatocita è una cellula epiteliale secretoria polare avente una membrana basolaterale (sinusoidale e laterale) e apicale (tubulare).
La formazione della bile include la cattura di acidi biliari e altri ioni organici e inorganici, il loro trasporto attraverso la membrana basale (sinusoidale), il citoplasma e la membrana tubulare. Questo processo è accompagnato dalla filtrazione osmotica dell'acqua contenuta negli epatociti e nello spazio paracellulare. L'identificazione e la caratterizzazione delle proteine di trasporto delle membrane sinusoidali e tubulari sono complesse. Particolarmente difficile è lo studio dell'apparato secretorio dei tubuli, tuttavia, ad oggi, una tecnica per ottenere epatociti raddoppiati in una coltura di breve durata è stata sviluppata e dimostrata affidabile in molti studi.La clonazione delle proteine di trasporto ci consente di caratterizzare separatamente la funzione di ciascuno di essi.
Il processo di formazione della bile dipende "dalla presenza di alcune proteine portatrici nelle membrane basolaterali e tubulari" Na +, K svolge il ruolo di forza motrice della secrezione + - ATPasi della membrana basolaterale, che fornisce un gradiente chimico e una differenza di potenziale tra l'epatocita e lo spazio circostante. Na +, K + - L'ATPasi scambia tre ioni di sodio intracellulare per due ioni di potassio extracellulare, mantenendo un gradiente di concentrazione di sodio (alto all'esterno, basso all'interno) e potassio (basso fuori, alto dentro). Di conseguenza, il contenuto cellulare ha una carica negativa (-35 mV) rispetto allo spazio extracellulare, che facilita la cattura di ioni caricati positivamente e l'escrezione di ioni con carica negativa. Na +, K + -ATPase non viene rilevato nella membrana tubolare. Il flusso della membrana può influenzare l'attività dell'enzima.
Cattura sulla superficie di una membrana sinusoidale
La membrana basale (sinusoidale) ha molti sistemi di trasporto per catturare anioni organici, la cui specificità di substrato si sovrappone parzialmente. La caratterizzazione delle proteine portatrici è stata precedentemente fornita sulla base dello studio delle cellule animali. La recente clonazione delle proteine di trasporto umano ha permesso di caratterizzare meglio la loro funzione. La proteina di trasporto per anioni organici (proteina di trasporto dell'anione organica - OATP) è indipendente dal sodio, trasporta molecole di un numero di composti, inclusi acidi biliari, bromsulfaleina e, probabilmente, bilirubina. Si ritiene che il trasporto della bilirubina negli epatociti sia effettuato anche da altri trasportatori. La cattura degli acidi biliari coniugati con taurina (o glicina) viene effettuata dalla proteina di trasporto sodio / taurocolato (proteina cotonosporante acido sodico / biliare - NTCP).
Il trasferimento di ioni attraverso la membrana basolaterale coinvolge una proteina che scambia Na + / H + e regola il pH all'interno della cellula. Questa funzione viene anche eseguita dalla proteina cotransport per Na + / HCO 3 -. Sulla superficie della membrana basolaterale vengono anche catturati solfati, acidi grassi non esterificati e cationi organici.
Trasporto intracellulare
Il trasporto di acidi biliari nell'epatocita viene effettuato utilizzando proteine citosoliche, tra le quali il ruolo principale è svolto dalla Z-idrossisteroide deidrogenasi. La glutatione-S-transferasi e le proteine leganti gli acidi grassi sono di minore importanza. Il reticolo endoplasmatico e l'apparato di Golgi sono coinvolti nel trasferimento degli acidi biliari. Apparentemente, il trasporto vescicolare viene attivato solo quando gli acidi biliari vengono significativamente iniettati nella cellula (a concentrazioni superiori a quelle fisiologiche).
Il trasporto di proteine in fase liquida e ligandi, come le IgA e le lipoproteine a bassa densità, viene effettuato mediante transcitosi vescicolare. Il tempo di trasferimento dalla membrana basolaterale a quella tubulare è di circa 10 minuti. Questo meccanismo è responsabile solo per una piccola parte della corrente totale della bile e dipende dallo stato dei microtubuli.
Secrezione tubulare
La membrana tubolare è una sezione specializzata della membrana plasmatica dell'epatocita contenente proteine di trasporto (principalmente ATP-dipendenti) responsabili del trasferimento di molecole in bile contro un gradiente di concentrazione. Enzimi come fosfatasi alcalina, GGTP sono anche localizzati nella membrana canalicolare. Il trasferimento di glucuronidi e glutatione-S-coniugati (ad esempio, la bilirubina diglucuronide) viene effettuato con l'aiuto di una proteina di trasporto multispecifica canalicolare per anioni organici (ghiandola cMOAT); trasportatore - SWAT), la cui funzione è parzialmente controllata dal potenziale intracellulare negativo. La corrente della bile, che non dipende dagli acidi biliari, è apparentemente determinata dal trasporto di glutaion, così come dalla secrezione canalicolare del bicarbonato, eventualmente con la partecipazione di una proteina che scambia Cl - / HCO 3 -.
Un ruolo importante nel trasporto di sostanze attraverso la membrana tubulare appartiene a due enzimi della famiglia della glicoproteina P; entrambi gli enzimi sono dipendenti dall'ATP. La proteina multidrug resistance protein 1 (MDR1) multidrug resistance 1 trasferisce cationi organici e rimuove anche i farmaci citostatici dalle cellule tumorali, causando la loro resistenza alla chemioterapia (da qui il nome protein). Il substrato endogeno di MDR1 è sconosciuto. MDR3 trasporta i fosfolipidi e agisce come un flippase per la fosfatidilcolina. La funzione di MDR3 e la sua importanza per la secrezione di fosfolipidi nella bile sono raffinate in esperimenti su topi privi della glicoproteina mdr2-P (analoga a MDR3 umana). In assenza di fosfolipidi nella bile, gli acidi biliari causano danni all'epitelio biliare, infiammazione dei dotti e fibrosi periduttolare.
L'acqua e gli ioni inorganici (specialmente il sodio) vengono escreti nei capillari biliari lungo il gradiente osmotico per diffusione attraverso contatti stretti semipermeabili a carica negativa.
La secrezione biliare è regolata da molti ormoni e messaggeri secondari, tra cui cAMP e protein chinasi C. L'aumento della concentrazione di calcio intracellulare inibisce la secrezione biliare. Il passaggio della bile attraverso i tubuli è dovuto a microfilamenti che forniscono motilità e contrazione dei tubuli.
Secrezione duttile
Le cellule epiteliali dei dotti distali producono un segreto ricco di bicarbonato che modifica la composizione della bile tubulare (il cosiddetto corrente duttulare, bile).Il processo di secrezione produce cAMP, alcune proteine di trasporto della membrana, inclusa una proteina che scambia Cl - / HCO 3 -, e regolatore della conduzione transmembrana nella fibrosi cistica -canale a membrana per Cl -, cAMP regolabile. La secrezione duttulare è stimolata dalla secretina.
Si presume che l'acido ursodesossicolico sia attivamente assorbito dalle cellule duttulari, scambiate con bicarbonati, riciclate nel fegato e successivamente espulse nuovamente nella bile ("shunt choleepatico"). Forse questo spiega l'effetto coleretico dell'acido ursodesossicolico, accompagnato da un'elevata secrezione di bicarbonato biliare nella cirrosi sperimentale.
La pressione nei dotti biliari, in cui si verifica la secrezione biliare, è normalmente di 15-25 cm di acqua. Art. Aumentare la pressione a 35 cm di acqua. Art. porta alla soppressione della secrezione biliare, allo sviluppo dell'ittero. La secrezione di bilirubina e acidi biliari può essere completamente interrotta, mentre la bile diventa incolore (galla bianca)e assomiglia al fluido mucoso.
La bile è un liquido plasmatico isosmotico costituito da acqua, elettroliti e sostanze organiche (acidi biliari, fosfolipidi, colesterolo, bilirubina). Gli acidi biliari (o loro sali) sono il principale componente organico della bile. Gli acidi biliari entrano nella bile da due fonti: (1) acidi biliari primari (colico e chenodeossicolico), che sono sintetizzati dal colesterolo nel fegato; (2) gli acidi biliari secondari (desossicolico, litocolico e ursodesossicolico) sono formati dall'azione dei batteri intestinali dagli acidi biliari primari. Gli stessi acidi biliari sono costituiti da due componenti importanti che determinano le loro proprietà fisiologiche e fisico-chimiche: (1) un nucleo steroideo con sostituenti idrossilici; (2) una catena laterale alifatica (Figura 7-4).
Fig. 7-4.
Gli acidi biliari sono costituiti da due componenti: un nucleo con estremità ossidrilica e una catena laterale alifatica. Nella figura, l'acido colico è mostrato come un esempio di acido 3-idrossi (3? -, 7? -, 12? -OH). Altri esempi sono acidi biliari contenenti desossicolato (3? -, 12? -OH), chenodeoxycholate (3? -, 7? -OH) e litocollo (3? -OH)
Nella maggior parte dei mammiferi, gli acidi biliari primari contengono da tre a sette sostituenti idrossilici, il cui numero influenza la loro solubilità in acqua (idrofilia). Poco dopo la formazione, gli acidi biliari primari subiscono modifiche nel gruppo carbossilico terminale. Ciò si verifica durante la fase epatica della circolazione enteroepatica degli acidi biliari secondari e la loro coniugazione con glicina o taurina. La presenza di componenti idrofilici (componenti ossidrilici e legami ammidici della catena laterale alifatica) e componenti idrofobi (nucleo steroideo) consente alle molecole di acido biliare coniugate di agire come un composto anfotero. Questo dà loro l'opportunità di formare micelle (aggregati polimecolari) al di sopra della concentrazione micellare critica. A loro volta, le molecole degli acidi biliari sono in grado di dissolvere altre sostanze anfotere (colesterolo, fosfolipidi) con la formazione di micelle miste. Questo ruolo detergente degli acidi biliari è importante per la stabilizzazione dello stato fisico-chimico della bile, la digestione e l'assorbimento dei grassi.
La sintesi degli acidi biliari dal colesterolo è regolata da un meccanismo di feedback negativo, anche se la natura della regolazione a livello molecolare e biochimico non è ancora completamente compresa. Microsomale 7? -Idrossilazione del colesterolo è un passo fondamentale nella sintesi degli acidi biliari. L'acido chetodesossicolico, usato per dissolvere i calcoli della colecisti, inibisce la sintesi degli acidi biliari e quindi aumenta i livelli di colesterolo nel sangue. Quando si utilizza l'acido ursodesossicolico, tali cambiamenti non vengono osservati anche con un trattamento a lungo termine.
La formazione della bile avviene su entrambe le superfici sinusoidali e tubolari della membrana epatocitaria ed è sia un processo intracellulare che paracellulare. In contrasto con la filtrazione glomerulare nei reni, che passivamente sotto l'azione delle forze idrostatiche, durante la formazione della bile, vi è un trasferimento attivo di componenti organici e inorganici nel lume dei tubuli e il trasporto passivo dell'acqua. Pertanto, i processi di secrezione biliare sono simili ai processi di secrezione negli acini del pancreas, l'epitelio dei tubuli renali. La formazione della bile tubolare può essere suddivisa in due tipi (Fig. 7-5): (1) formazione di bile, a seconda della secrezione di acidi biliari, definita come il rapporto tra la quantità di bile secreta nei tubuli e la quantità di sali biliari secreti; (2) formazione di bile, indipendente dalla secrezione di acido biliare, che può essere rappresentata come una secrezione attiva di elettroliti inorganici e altre sostanze e si riflette sul grafico come un'intersezione y di questa linea. In altre parole, la formazione di bile associata alla secrezione acida è la velocità di flusso della bile, a seconda della presenza di sali biliari osmoticamente attivi nei dotti biliari e della formazione di bile non correlata alla secrezione acida in assenza di sali biliari. Il rapporto tra il tasso di formazione della bile e la formazione di sali di acidi biliari è non lineare con piccole quantità di bile secreta e non può corrispondere alla relazione lineare mostrata in Fig. 7-5. Pertanto, entrambi i tipi di formazione biliare dovrebbero essere considerati come indicatori interconnessi della formazione della bile.
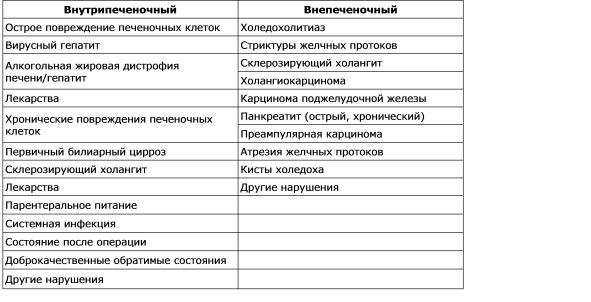
La violazione della formazione della bile è chiamata colestasi. La sequenza delle manifestazioni patologiche, fisiologiche e cliniche emergenti della colestasi dipende dalla causa di essa. Uno studio morfologico del materiale della bile dell'epatobiopsia viene rilevato nei tubuli degli epatociti pericentral, si nota la dilatazione dei tubuli e lo studio dell'ultrastruttura rivela una diminuzione del numero di microvilli. La colestasi può essere definita come un difetto funzionale nella formazione della bile a livello degli epatociti (colestasi intraepatica), così come la secrezione organica o meccanica e il deflusso della bile (colestasi extraepatica). Le cause più comuni di colestasi intraepatica ed extraepatica sono riportate in Tabella. 7-2. Esistono diversi meccanismi che giocano un ruolo importante nella patogenesi della colestasi intraepatica: compromissione e danneggiamento della funzione della membrana sinusoidale; interruzione delle funzioni degli organelli intracellulari degli epatociti; danno e rottura della membrana tubolare. Quindi, non esiste un singolo meccanismo per la colestasi in varie situazioni cliniche, e la molteplicità dei meccanismi può portare a vari disturbi. Clinicamente, la colestasi è caratterizzata da un aumento dei livelli ematici di molte sostanze, tra cui la bilirubina, i sali biliari, il colesterolo, che sono normalmente secreti nella bile. Nell'analisi biochimica del sangue con colestasi, si osserva un aumento sproporzionato dell'attività della fosfatasi alcalina e della concentrazione di bilirubina in parallelo con i cambiamenti nel livello di aminotransferasi, che verranno discussi di seguito.
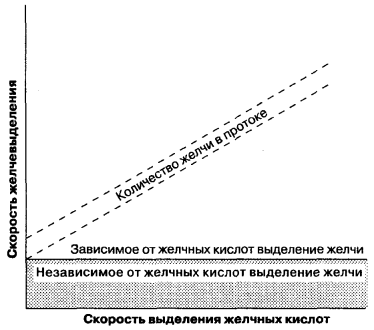
Fig. 7-5.
Formazione di bile associata alla secrezione di acido biliare, formazione di bile indipendente dalla secrezione di acido biliare. (Da: Moseley R. H., secrezione di Bile. In: Yamada T., Alpers D. H., Owyang C., Powell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995: 387.)
Tabella 7 - 2.
Nel fegato si forma il succo digestivo più importante: la bile.
La bile è prodotta dagli epatociti mediante trasporto attivo e passivo di acqua, colesterolo, bilirubina, cationi in essi. Negli epatociti del colesterolo si formano acidi biliari primari - colici e desossicolici. Un complesso idrosolubile è sintetizzato dalla bilirubina e dall'acido glucuronico. Entrano nei capillari e nei dotti biliari, dove gli acidi biliari si combinano con glicina e taurina. Di conseguenza, si formano gli acidi glicoclorico e taurocolico. Il bicarbonato di sodio è formato dagli stessi meccanismi del pancreas.
La bile è prodotta dal fegato tutto il tempo. Nel suo giorno si forma circa 1 litro. Gli epatociti espellono la bile primaria o epatica. Questo liquido è una reazione alcalina giallo-oro. Il suo pH = 7.4 - 8.6. Consiste del 97,5% di acqua e del 2,5% di solidi. Il residuo secco contiene:
1. Sostanze minerali Sodio, potassio, cationi di calcio, bicarbonato, anioni fosfato, anioni di cloro.
2. Acidi acidi - taurocholic e glycocholic.
3. Pigmenti biliari - bilirubina e sua forma ossidata biliverdina. La bilirubina dà il colore della bile.
4. Colesterolo e acidi grassi.
5. Urea, acido urico, creatinina.
Dal di fuori del sistema digestivo, lo sfintere di Oddi, situato alla foce del dotto biliare comune, è chiuso, la bile escreta si accumula in cistifellea. Qui l'acqua viene riassorbita da esso e il contenuto di componenti organici di base e di mucina aumenta di 5-10 volte. Pertanto, la bile cistica contiene il 92% di acqua e l'8% di residuo secco. È più scuro, più spesso e più viscoso del fegato. A causa di questa concentrazione, la vescica può accumulare la bile per 12 ore. Durante la digestione, lo sfintere di Oddi e lo sfintere di Lutkens nel collo della vescica si aprono. La bile entra nel duodeno.
Bile Value:
1. Gli acidi malici emulsionano una parte dei grassi, trasformando grosse particelle di grasso in goccioline fini.
2. Attiva gli enzimi del succo intestinale e pancreatico, in particolare la lipasi.
3. In combinazione con gli acidi biliari, l'assorbimento degli acidi grassi a catena lunga e delle vitamine liposolubili avviene attraverso la membrana degli enterociti.
4. Zhelchka promuove la risintesi dei trigliceridi negli enterociti.
5. Attiva le pepsine e neutralizza anche l'acido chimo proveniente dallo stomaco. Questo assicura il passaggio dalla digestione gastrica a quella intestinale.
6. Simula la secrezione di succhi pancreatici e intestinali, nonché la proliferazione e la desquamazione degli enterociti.
7. Rafforza la motilità intestinale.
8. Ha un effetto batteriostatico sui microrganismi intestinali e quindi impedisce lo sviluppo di processi putrefattivi in esso.
La regolazione della formazione della bile e dell'escrezione biliare è principalmente effettuata da meccanismi umorali, anche se quelli nervosi svolgono un certo ruolo. Lo stimolatore più potente della formazione di bile nel fegato è gli acidi biliari, assorbiti nel sangue dall'intestino. Viene anche potenziato dalla secretina, che contribuisce ad aumentare il bicarbonato di sodio nella bile. Il nervo vago stimola la produzione di bile, l'inibizione simpatica.
Quando il chimo entra nel duodeno, le cellule I iniziano a rilasciare le sue cellule del colecistochinina-pancreozil. Soprattutto questo processo è stimolato da grassi, tuorlo d'uovo e solfato di magnesio. CCK-PZ rafforza le contrazioni dei muscoli lisci della vescica, i dotti biliari, ma rilassa gli sfinteri di Lutkens e Oddi. La bile viene rilasciata nell'intestino. I meccanismi dei riflessi giocano un piccolo ruolo. Chyme irrita i chemocettori dell'intestino tenue. Gli impulsi da loro entrano nel centro digestivo del midollo allungato. Da lui sono sul vago al tratto biliare. Gli sfinteri si rilassano e i muscoli lisci del contratto vescicale. Promuove l'escrezione biliare.
Le malattie più gravi sono l'epatite e la cirrosi. Molto spesso l'epatite è il risultato di un'infezione (epatite infettiva A, B, C) e dell'esposizione a prodotti tossici (alcol). Nell'epatite, gli epatociti sono colpiti e tutte le funzioni epatiche sono compromesse. La cirrosi è il risultato dell'epatite. La violazione più comune dell'escrezione biliare è la colelitiasi. La maggior parte dei calcoli biliari è formata dal colesterolo, dal momento che la bile di tali pazienti è supersatura con essi.
Il fegato è il più grande organo del corpo ed è centrale per il metabolismo. Svolge molte funzioni, partecipando al metabolismo di proteine, carboidrati, grassi, ormoni e vitamine, nonché alla neutralizzazione di molte sostanze endogene ed esogene. Questi processi sono descritti nei libri di testo di chimica fisiologica e in questa sezione in relazione a sistema digestivo considereremo solo funzione epatica escretoriavale a dire secrezione di bile. bile consiste di acqua, sali minerali, muco, lipidi di colesterolo e lecitina e due tipi di componenti specifici - acidi biliari e pigmento di bilirubina. Gli acidi biliari sono detergenti e la loro azione emulsionante gioca un ruolo importante nella digestione dei lipidi. La bilirubina è il prodotto finale della scomposizione dell'emoglobina da eliminare dal corpo.
^
Formazione di bile
Anatomia funzionaleCellule del fegato (Epatociti)formare un piatto nello spessore di una cella, separato da fenditure strette (Spazio Disse)che rappresenta il completato
^ 766 PARTE VIII. CIBO, DIGESTIONE ED ESTRATTO
Dal sangue sinusoidaleequivalente a capillari. Nelle pareti delle sinusoidi ci sono pori attraverso i quali possono passare macromolecole così grandi come l'albumina e le lipoproteine. Nel più piccolo kanaltsam-capillari biliari delimitati dalle membrane plasmatiche di due epatociti adiacenti - la bile viene raccolta in dimensioni maggiori i canaliculi di Goring,le cui pareti, come le più grandi tubulo di intercollegamentoe dotti biliariformato da cellule secretorie cubiche. I tubuli piccoli all'interno dei lobuli di fegato e tra di loro si fondono in quelli più grandi, formando alla fine dotto epatico.Da questo dotto parte dotto cisticoalla cistifellea. Dopo la fusione, si formano i dotti epatici e cistici dotto biliare comuneapertura nel duodeno nella parte superiore della papilla del Vater, dietro o vicino al dotto pancreatico (Fig. 29.1).
^ Funzioni della bile.Bile svolge molte importanti funzioni. Con lei vengono visualizzati i prodotti finaliscambio, come la bilirubina, così come i farmaci e le tossine. assegnazione conbile colesterolosvolge un ruolo importante nella regolazione del suo equilibrio. Acidi biliariessenziale per l'emulsione e l'assorbimento dei grassi. Inoltre, la bile contiene acqua, sali minerali e muco. Circa 24 ore al giorno 600 mlbile e 2/3 di questa quantità provengono dai tubuli e 1/3 dei condotti più grandi.
^ Bile tubolare si forma in quantità approssimativamente uguali con la partecipazione di due diversi meccanismi, bile-dipendenti e bile-dipendenti (figura 29.27).
^ Secrezione dipendente da acido biliare. C'è una stretta relazione tra portata di bilee secrezione di acido biliare.Nella bile canaliculare, la concentrazione di acidi biliari è 100 volte superiore a quella del sangue portale, pertanto sono considerati secreti da trasporto attivo che coinvolge il vettore.Seguendo gli acidi biliari lungo il gradiente osmotico, l'acqua scorre nei tubuli, quindi la bile è isotonica con il sangue.
Ci sono due fonti di acidi biliari. In primo luogo, sono sintetizzati de novo dal colesterolo negli epatociti stessi con la partecipazione di 7-idrossilasi. Questo enzima svolge un ruolo chiave e controlla la velocità di sintesi degli acidi biliari attraverso un meccanismo di feedback. In secondo luogo, gli epatociti sono in grado di assorbire attivamente gli acidi biliari sangue portaleed espellili in tubuli (vedi anche figura 29.29). Questa estrazione è molto efficace; con un singolo passaggio di sangue attraverso il fegato, l'80% degli acidi biliari viene estratto da esso. pertanto
La concentrazione di acidi biliari nel sangue periferico è molto inferiore rispetto al sistema portale. Poiché gli acidi biliari vengono estratti dal sangue 6 volte più velocemente di quanto non entrino nei tubuli, è quest'ultimo processo che limita la velocità della secrezione di acido biliare.
^ Secrezione indipendente dagli acidi biliari. il
Questo processo coinvolge ioni Na +, Cl -, HCO 3 - e acqua. La forza trainante è trasporto attivo Να + , possibilmente con bicarbonato. Una secrezione indipendente dagli acidi biliari stimola, in particolare, secretina.
Oltre agli acidi biliari, la bilirubina, il colesterolo e i fosfolipidi (principalmente lecitina) sono attivamente secreti nei canalicoli (Figura 29.27). Insolubile in acqua ("Indiretta") bilirubina,la maggior parte dei quali è formata da emoglobina di globuli rossi invecchiati, entra negli epatociti sotto forma di un aggregato colloidale associato all'albumina. La sua formazione giornaliera è di circa 4 g / kg di peso corporeo o 200-300 mg / die. Negli epatociti, l'80% della bilirubina è coniugata. conacido glucuronico e una piccola quantità di esso con acido solforico. In tale coniugato
^ CAPITOLO 29. FUNZIONI DEL TRATTO GASTROINTESTINALE 767
La bilirubina è escreta nella bile. (Bilirubina "diretta").In generale, i farmaci e le tossine vengono rimossi allo stesso modo.
^ Modifica della bile nei dotti biliari
(fig 29.27). Nei condotti in cui i tubuli si aprono, la bile primaria subisce delle modifiche. Questo processo assomiglia al processo di modifica del filtrato glomerulare nei tubuli renali (p.785), e in modo simile è calcolato autorizzazionesolo nel caso della bile viene usata una sostanza inerte anziché inulina eritritoloo mannitolo,che sono secreti nei tubuli, ma non riassorbiti. Studi simili hanno dimostrato che circa 180 ml di bile, o 1/3 della sua quantità totale, vengono secreti nei dotti con secrezione attiva di NSO ^ ". Questo processo è stimolato secretina.
^
Bile epatica e cistifellea
La composizione della bile epatica(tab. 29.3). La bile secreta dal fegato ad una velocità di 0,4 ml / min ha un colore dorato, che si spiega con la presenza di bilirubina in esso. La concentrazione di elettroliti in questa bile è la stessa del plasma, tranne per il fatto che contiene il doppio di HCO3 "e un po 'meno di CI ~. Allo stesso tempo, la composizione delle sostanze organiche bile è molto diversa dal plasma, dato che nella bile sono rappresentato quasi esclusivamente da acidi biliari, colesterolo e fosfolipidi.
 |
^ Acidi biliarisono formati nel fegato da colesterolo come risultato della sua idrossilazione e l'aggiunta di un gruppo carbossilico. Gli acidi che si formano nel fegato sono acidi biliari primari;questi includono chenodeossicolico(dioxyform) e colico(Trioksiforma) l'acido.Nel fegato, non sono in forma libera, ma sotto forma di coniugati con glicina e taurina, e i coniugati con glicina si formano tre volte di più, poiché la quantità di taurina è limitata. Gli acidi biliari coniugati sono più solubili in acqua,
Che non sono coniugati e hanno una maggiore capacità di dissociarsi e formare sali biliari con cationi, principalmente con ioni Na +. In un ambiente acido (pH 4.0), i sali di acidi biliari sono insolubili e precipitati, ma a valori di pH fisiologici (nell'intestino tenue), sono ben solubili.
Nella parte distale dell'ileo e nel colon, alcuni dei sali degli acidi biliari primari subiscono la deidrossilazione sotto l'azione di batteri anaerobici e si trasformano in bile acido-litochico secondario(monoossiforme) e deoxycholic(Dioksiforma). Gli acidi chenossossicolico, colico e desossicolico sono presenti in un rapporto 2: 2: 1. L'acido litocholico è presente solo in alcune frazioni, poiché la maggior parte di questo è escreto.
L'effetto emulsionante degli acidi biliari sui grassi si basa principalmente sulla loro capacità di formare micelle.Le molecole di acido biliare hanno una struttura tridimensionale in cui i gruppi idrossilici carbossile e idrossile si trovano sullo stesso lato della molecola e la parte idrofobica della molecola (nucleo steroideo, gruppi metilici) si trova sul lato opposto, a causa della quale le molecole di acido biliare hanno e idrofilo,e proprietà lipofile.A causa di questa struttura, le molecole degli acidi biliari agiscono come detergenti: all'interfaccia delle fasi lipidica e acquosa, formano un film quasi monomolecolare in cui i gruppi idrofili vengono trasformati in fase acquosa e lipofila in fase lipidica. Nella fase acquosa, gli acidi biliari formano aggregati ordinati. -mitsellya condizione che la loro concentrazione raggiunga un certo livello, chiamato concentrazione micellare critica(1-2 mmol / l). La regione interna, lipofila della micella può contenere lipidi,per esempio, colesterolo e fosfolipidi; tali micelle sono chiamate miste (Fig. 29.28). Il colesterolo stesso è insolubile in acqua, ma può essere in soluzione nella composizione delle micelle. Se la sua concentrazione supera la capacità delle micelle, forma un precipitato cristallino; questo processo è alla base della formazione di calcoli biliari di colesterolo (p.769).
^ La composizione della bile cistica (tab. 29.3). La capacità della cistifellea è di soli 50-60 ml. Allo stesso tempo, il fegato secerne la bile ad una velocità di 600 ml / giorno, e metà di questa quantità passa attraverso la cistifellea prima di entrare nell'intestino tenue. La differenza tra il volume della bile che entra nella cistifellea e la sua capacità viene compensato riassorbimento altamente efficienteacqua nella cistifellea. Entro poche ore, il 90% dell'acqua può essere riassorbita dalla bile. a
^
768 PARTE VIII. CIBO, DIGESTIONE ED ESTRATTO
Questa materia organica rimane nella cistifellea e la loro concentrazione nella bile aumenta. La forza trainante del riassorbimento è trasporto attivo di ioni Nα *con la partecipazione di una "pompa" incorporata nelle membrane basali e laterali delle cellule e attivata da (Na + -K +) - ATPasi. Seguendo gli ioni Na +, gli ioni Cl - e HCO 3 si diffondono nella direzione del gradiente elettrico o trasportati dai trasportatori. Come risultato del riassorbimento di HCO 3 - il pH della colecisti diminuisce a 6,5 vs 8,2 nella bile epatica. Come risultato della creazione di un'alta concentrazione di ioni Na + nell'epitelio della cistifellea nello spazio extracellulare, si verifica un gradiente osmotico che porta al pompaggio di acqua, che quindi fluisce nei capillari (pagina 751).
^ Cistifellea motilità. Nello stato di digiuno, la bile si accumula nella cistifellea e in
Il tempo di mangiare viene assegnato come risultato delle contrazioni della cistifellea. Lo stimolatore principale dell'attività contrattile della cistifellea è colecistochinina,secreto dalla membrana mucosa del duodeno dopo l'ammissione ad esso di grasso contenente chimo. Le contrazioni della cistifellea sono anche stimolate in una certa misura. nervo errantee parasimpaticolitici.Iniziano entro 2 minuti dopo che il cibo grasso entra in contatto con la mucosa intestinale e dopo 15-90 minuti la vescica viene completamente svuotata. La motilità della cistifellea coinvolge due processi. Inizialmente, si sviluppa una contrazione tonica, a seguito della quale il diametro della cistifellea diminuisce, e quindi vengono imposte contrazioni periodiche su questo effetto, la cui frequenza è 2-6 / min. Come risultato di questi due processi, viene creata una pressione di 25-30 mm Hg. Art.
^
Circolazione enteroepatica
Circolazione di acido biliare(fig 29.29). Gli acidi biliari sono secreti nel duodeno come micelle miste. Nonostante la diluizione degli acidi biliari dal contenuto dello stomaco, la loro concentrazione nell'intestino è di circa 10 mmol / l e rimane al di sopra della concentrazione critica della formazione di micelle. Qui, oltre al colesterolo e alla lecitina, le micelle includono i prodotti della rottura del grasso idrolitico - acidi grassie monogliceridi.Al primo contatto delle micelle con la parete intestinale
^
CAPITOLO 29. FUNZIONI DEL TRATTO GASTROINTESTINALE 769
I lipidi si diffondono attraverso la membrana del bordo del pennello in enterociti e gli acidi biliari rimangono nel lume intestinale, ma con un ulteriore passaggio attraverso l'intestino, gli acidi biliari vengono assorbiti dal trasporto attivo e passivo.
Circa il 50% degli acidi biliari viene assorbito passivamente nell'intestino. Come risultato della scissione dei coniugati di acido biliare e della deidrossilazione di questi ultimi dall'azione dei batteri intestinali, la loro solubilità nei lipidi aumenta e la diffusione passiva è facilitata.
^ Aspirazione attiva gli acidi biliari si verificano esclusivamente nell'ileo terminale - un fenomeno raro, noto solo per l'assorbimento di vitamina B 12. Solo quegli acidi biliari che hanno un'alta polarità che impediscono il loro assorbimento passivo, come i coniugati taurini, sono soggetti ad assorbimento attivo. Il processo di assorbimento degli acidi biliari nell'ileo terminale è caratterizzato da segni tipici di trasporto attivo: cinetica di saturazione e inibizione competitiva. Una piccola quantità di acidi biliari (7-20%) non è inclusa nell'assorbimento attivo o passivo ed è eliminata dal corpo.
La presenza di acidi biliari nel colon sembra giocare un ruolo importante nella regolazione della consistenza delle feci. Quando la concentrazione di Dioxy Acid nel colon supera i 3 mmol / l, una quantità significativa di elettroliti e acqua viene secreta nel lume intestinale, che porta alla diarrea. Forma pronunciata di questo Diarrea "Hologennogo"può essere osservato con resezione o malattia dell'ileo terminale e per il suo trattamento utilizzando il legame degli acidi biliari utilizzando lo scambiatore di ioni colestiramina.
Quando ingerito acidi biliari assorbiti al fegatoi coniugati sono appena formati e alcuni acidi biliari secondari subiscono l'idrossilazione. La perdita di acidi biliari nelle feci (0,2-0,6 g / die) è compensata dalla loro sintesi.
^ Pool totale di acidi biliari nel corpo è di circa 3,0 g Questa quantità non è sufficiente per garantire la lipolisi dopo un pasto; in particolare, il consumo di cibi grassi richiede 5 volte più acidi biliari. Tuttavia, il corpo non è carente di acidi biliari, poiché circolano molte volte attraverso l'intestino e il fegato. (circolazione enteroepatica).La frequenza con cui il pool di acidi biliari fa un ciclo completo dipende dal regime alimentare e varia da 4 a 12 cicli al giorno.
^ Circolazione della bilirubina. Pigmento gallo bilirubina,come acidi biliari e lipidi, entra nell'intestino sotto forma di glzhuronid. Solo una piccola quantità di questo composto polare
Riassorbito nella cistifellea e nell'intestino tenue. Nell'ileo terminale e (principalmente) nel colon, il coniugato di bilirubina viene scisso dall'azione delle idrolasi batteriche. Nello stesso tempo la bilirubina si trasforma in urobilinogeno,che, insieme ad altri prodotti di decomposizione della bilirubina, conferisce colore marrone alle feci. Meno del 20% dell'urobilinogeno viene riassorbito e di questa quantità circa il 90% rientra nel fegato e ritorna alla bile, mentre il restante 10% viene escreto nelle urine.
Aspetti fisiopatologici. Livelli elevati di urobilinogeno nelle urine possono indicare malattia del fegato,accompagnato da una violazione dell'escrezione di bilirubina. La completa assenza di urobilinogeno nelle urine, il colore chiaro delle feci e il colore giallo indicano blocco completo della cistifellea;in questa situazione, la bilirubina non entra affatto nell'intestino e l'urobilinogeno non si forma.
La violazione più nota e diffusa della fisiologia normale del sistema biliare è la precipitazione del colesterolo con la formazione di calcoli biliari di colesterolo. Il colesterolo, come la lecitina, è in uno stato di dissoluzione solo come parte di micelle miste. Se la concentrazione aumenta xo.jecmepo.ia o la concentrazione di acidi biliari o lecitina diventa inferiore ai livelli critici,il colesterolo precipita. Tra i fattori che causano un aumento del contenuto di colesterolo relativo sono gli estrogeni, una dieta di carboidrati, sovrappeso e processi che riducono la concentrazione di acidi biliari, come l'infiammazione dell'ileo. (Malattia di Crohn)o resezione. In alcuni casi, la supplementazione di acido biliare orale può essere sufficiente per trasformare la litio litogenea in alitogenico, in cui i calcoli di colesterolo possono dissolversi. Gli acidi cinodoossicolico e urodesossicolico sono i più adatti a questo scopo, poiché non causano la diarrea.
La manifestazione clinica del metabolismo compromesso della bilirubina è l'ittero. La pelle gialla è associata ad elevati livelli plasmatici di bilirubina, che possono verificarsi nei seguenti casi:
con aumento della formazione di bilirubina a seguito di un aumentato collasso degli eritrociti (ittero emolitico);
come risultato di interruzione del processo di coniugazione o trasporto di bilirubina negli epatociti, come, per esempio, ittero gravidoo l'ittero di maternità di Gilbert;
con deflusso ritardato della bile, ad esempio a causa di calcoli biliari o tumori localizzati nella regione del dotto biliare (ittero ostruttivo).