Formiranje žuči. Fiziologija formiranja žuči. Bile formation
Nastajanje žuči javlja se u jetri kao rezultat aktivnog i pasivnog transporta supstanci (voda, glukoza, kreatinin, elektroliti, hormoni, vitamini, itd.) Koji cirkuliraju u krvi, kroz ćelije i kontakte ćelija - stanica, kao i aktivna sekrecija komponenti žuči (žučnih kiselina) hepatocitima i reapsorpciju vode i određenih supstanci iz malih žučnih puteva i žučnih kamenaca. Fiziološki značaj ovog procesa je raznolik. Žuči se prvenstveno smatraju probavnom tajnom, jer žučne kiseline (uglavnom njihovi organski anioni) igraju ključnu ulogu u apsorpciji masti. Žuči emulgiraju masti, povećavajući površinu na kojoj se hidroliziraju pod utjecajem lipaze. Pod dejstvom žuči je rastvaranje proizvoda masne hidrolize. Promoviše njihovu apsorpciju i resintezu triglicerida u enterocitima. Žuči povećavaju aktivnost enzima pankreasa i creva (posebno lipaza), pojačavaju hidrolizu i apsorpciju proteina i ugljenih hidrata.
U suprotnosti sa varenjem masti slabo se probavljaju i druge prehrambene supstance, jer masnoća obavija najmanje čestice hrane i sprečava djelovanje enzima na njih. U takvim uslovima aktivnost crevnih bakterija dovodi do povećanih procesa raspadanja, fermentacije i formiranja gasa.
Žučni inherentni i regulatorni efekti - stimulacija formiranja žuči, izlučivanje žuči, motorička i sekretorna aktivnost tankog crijeva, kao i proliferacija i desquamation enterocita. Žuči inhibira proces probave želuca neutralizacijom kiseline koja je ušla u duodenum, i inaktivacijom pepsina, pripremom za varenje u crevima. Takođe ima regulatorni uticaj na evakuacionu aktivnost gastroduodenalnog kompleksa. Uloga žuči u apsorpciji vitamina topljivih u mastima (A, D, E i K), holesterola, soli kalcijuma iz crijevnog lumena je važna.
Pored toga, formiranje i izlučivanje žuči smatra se neobičnim načinom izlučivanja određenih molekula i jona koji se ne mogu izlučivati kroz bubrege. Među njima, najznačajniji su holesterol (izveden kao slobodni holesterol, njegovi esteri i žučne kiseline), bilirubin, kao i molekule bakra i gvožđa. Zbog toga se žuč smatra izlučujućom tekućinom.
Žuči se sastoje od 80% vode i 20% otopljenih supstanci. Ovo uključuje žučne kiseline i njihove soli (oko 65%), žučne fosfolipide (oko 20%, uglavnom zbog lecitina), proteine (oko 5%), holesterol (4%), konjugirani bilirubin (0,3%), enzime , imunoglobulini, kao i brojne egzogene i endogene supstance koje se luče žuči (biljni stiren, vitamini, hormoni, lekovi, toksini, metalni joni - bakar, gvožđe, kalijum, natrijum, kalcijum, cink, magnezijum, živa, itd). U proseku se dnevno izlučuje 600-700 ml žuči (od 250 do 1500 ml, približno 10,5 ml na 1 kg tjelesne težine). Istovremeno, oko 500 ml / dan ukupne zapremine obezbeđuje se izlučivanjem hepatocita i oko 150 ml / dan ćelija žučnih kanala izlučuje.
Bile formation (choleresis) ide kontinuirano, i protok žuči u duodenum (holekineza) događa se periodično. Na prazan želudac žuč ulazi u crevo u skladu sa periodičnom gladnom aktivnošću. Tokom perioda odmora, ide na SB, gdje se koncentrira, donekle mijenja svoj sastav i deponira se. Pored vode i soli, apsorbuju se i holesterol i slobodne masne kiseline. U tom smislu, razlikovati žuči jetre i žučne kese.
Žuči imaju malu enzimsku aktivnost; PH jetre je 7,3-8,0. Za razliku od intestinalnog sadržaja, ne sadrži gotovo nikakve bakterije. Čimbenici koji osiguravaju sterilnost žuči uključuju prisustvo žučnih kiselina (bakteriostatski učinak), bogat sadržaj imunoglobulina, izlučivanje sluzi, relativno siromaštvo supstrata žučne energije za bakterije.
Žuči su micelarne otopine. Holesterol, praktično nerastvorljiv u vodi, prenosi se u rastvorenom stanju u žuči zbog svoje micelarne strukture. Ovaj proces se naziva koloidno rastvaranje - solubilizacija.
Žučne kiseline su površinski aktivni, amfipatski (i hidrofobni i hidrofilni) molekuli sposobni za samo-agregaciju. U ovom slučaju, zahvaljujući vrlo uskoj koncentraciji, koja se naziva kritična koncentracija micelizacije, formiraju se jednostavne micele. Jednostavne micele imaju izraženu sposobnost da rastvaraju lipide, formirajući mešovite micele.
Vjeruje se da mješovite micele imaju cilindričnu strukturu: cilindrično stablo je ispunjeno polarnim lipidima, a molekuli žučne kiseline su smješteni između polarnih krajeva lipidnih molekula s hidrofilnim stranama okrenutim vodenoj sredini, koje određuju njihovu hidrofilnost (topljivost u vodi).
Mešane micele sadrže esencijalne komponente - žučne kiseline koje se nalaze van, fosfolipidi (uglavnom fosfatidilholin - lecitin) i holesterol, koji se nalaze unutar micele.
Boja žuči je žućkasto-smeđa zbog prisustva bilirubina, čiji je najveći dio predstavljen u obliku diglukuronida bilirubina, manji - u obliku monoglukuronid bilirubina. Povezani bilirubin nije dio micela.
Pošto je međustanično bilijarno drvo propusno za vodu, žučna kesa i žuč su izotonične.
Bile formation
Biliary secretion
Žučne kiseline su glavna komponenta bilijarne sekrecije, formiraju se samo u jetri. Primarne žučne kiseline - trihidroksiholni (holij) i dihidroksiholni (chenodeoksiholni) sintetišu se u hepatocitima holesterola. Sekundarne žučne kiseline (deoksikolični i u malim količinama - litoholni) formiraju se u debelom crevu od primarne kao rezultat bakterijske modifikacije nuklearnih hidroksilnih grupa (7a-dehidroksilacija pod kontrolom anaerobnih bakterija). Tercijarne žučne kiseline (uglavnom ursodeoksiholni) nastaju u jetri izomerizacijom sekundarnih žučnih kiselina.
Sinteza žučnih kiselina iz holesterola reguliše se negativnim povratnim mehanizmom: smanjeni povrat žučnih kiselina u hepatocite sa portalnom krvi dovodi do povećanja njihove biosinteze; bilo koje povećanje sinteze žučnih kiselina je praćeno odgovarajućim povećanjem formiranja holesterola.
Žučne kiseline su konjugirane u jetri sa aminokiselinama glicinom (oko 80%) ili taurinom (oko 20%). Kada se konzumira hrana bogata ugljikohidratima, količina glikoholnih kiselina se povećava u žuči, a kod visoko-proteinskih dijeta, taurokolne kiseline. Vezivanje aminokiselina sprečava apsorpciju žučnih kiselina u bilijarnom traktu i početnim delovima tankog creva (samo u terminalnom ileumu i debelom crevu). Pod djelovanjem bakterija moguća je hidroliza žučnih soli s formiranjem žučnih kiselina, glicina ili taurina.
Nakon biosinteze žučnih kiselina, kao što je gore navedeno, njihova karboksilna grupa je vezana za amino grupu glicina ili taurina. Sa hemijske tačke gledišta, takvo vezivanje pretvara slabu kiselinu u jaču. Vezane žučne kiseline su najviše rastvorljive pri niskom pH i otpornije su na taloženje kalcijevim ionima (Ca 2+) u odnosu na nevezane žučne kiseline. Sa biološke tačke gledišta, proces vezivanja čini žučne kiseline nesposobnim za prodiranje kroz ćelijske membrane. Stoga se apsorbiraju u žučnim kanalima ili tankom crijevu, bilo u prisustvu molekula nosača, ili ako se žučne kiseline podvrgnu bakterijskom cijepanju. Jedini izuzetak od ovog pravila su žučne kiseline povezane sa glicin dihidroksi, budući da se mogu pasivno apsorbovati ako dobiju vodonični ion (H +).
Većina žučnih kiselina (oko 85-90%), primljenih sa protokom žuči u tanko crevo, apsorbuju se u krv. Međutim, većina povezanih žučnih kiselina koje se luče u tanko crevo se apsorbiraju netaknute. Manji dio kiseline se ne apsorbira netaknut, jer se podvrgava bakterijskom cijepanju u distalnom tankom crijevu. Oni se pasivno apsorbuju i, vraćajući se u jetru, ponovo se vežu i luče u žuč. Preostalih 10-15% žučnih kiselina se eliminira iz tijela uglavnom s izmetom. Ovaj gubitak žučnih kiselina se kompenzira njihovom sintezom u hepatocitima.
Ovaj proces crijevnog cijepanja i reapsorpcije u jetri je normalan dio metabolizma žučnih kiselina. Mali dio žučnih kiselina, koji se ne apsorbiraju, ulazi u debelo crijevo. Ovdje završava proces cijepanja. Pored toga, sekundarne žučne kiseline nastaju u kolonu pod dejstvom anaerobnih bakterija (vidi gore).
Deoksiholne i litoholne kiseline se delimično apsorbuju u debelom crevu i ponovo ulaze u jetru. Nakon povratka, metabolizam ovih žučnih kiselina je različit. Deoksiholna kiselina se vezuje za glicin ili taurin i cirkuliše sa primarnim žučnim kiselinama. Treba napomenuti da je kod starijih osoba deoksiholna kiselina glavna žučna kiselina u sastavu žuči. Litoholna kiselina se ne vezuje samo za glicin ili taurin, već dodatno za sulfat na C-3 položaju. Takvo "dvostruko" vezanje smanjuje mogućnost apsorpcije u crevima, zbog čega se litokolna kiselina brzo gubi iz sastava cirkulišućih žučnih kiselina - sadržaj žuči rijetko prelazi 5%. Većina (95%) žučnih kiselina, koje su deo žuči odraslih, su holne, ceneodoksiholne i deoksiholne kiseline.
Proces vezivanja žučnih kiselina u hepatocitima je veoma efikasan, tako da su žučne kiseline u potpunosti prisutne u konjugiranoj formi. Naknadno bakterijsko cijepanje i dehidroksilacija uzrokuju da žučne kiseline u fecesu budu u nevezanom stanju.
Gore opisani proces dehidroksilacije primarnih žučnih kiselina smanjuje njihovu sposobnost otapanja u vodi. Formiranje sekundarnih žučnih kiselina kod ljudi praktično nema fiziološki značaj, za razliku od životinja. Prekomerna apsorpcija deoksikolne kiseline u debelom crevu povećava rizik od holesterola u žuči. Litoholna kiselina se smatra hepatotoksičnim. U pokusima na životinjama, pokazalo se da nakupljanje litokolne kiseline u jetri dovodi do njenog poraza. Međutim, još nije dokazano da povećanje apsorpcije ove kiseline kod ljudi dovodi do poremećaja jetre.
Ursodeoksiholna kiselina, kao i deoksiholna kiselina, vezuje se u jetri i cirkuliše sa primarnim žučnim kiselinama. Međutim, metabolički put ove kiseline je veoma kratak i sadržaj vezanog ursodeoksiholne kiseline u žuči nikada ne prelazi 5% ukupne količine žučnih kiselina. Smatra se da formiranje ursodeoksiholne kiseline nema značajan fiziološki značaj.
Žučne kiseline su snažni rastvarači masti, stoga su citotoksični u koncentracijama koje dosežu kritičnu koncentraciju micelizacije. Istovremeno, vezana chenodeoxycholic i deoksiholne kiseline prisutne u žuči su više otrovne nego one povezane s holnim i ursodeoksiholnim. Iako su žučne kiseline dokazno citotoksične in vitro, epitel žučnih puteva i tankog creva se nikada ne oštećuje kao rezultat njihove visoke koncentracije. To je povezano, prije svega, s prisutnošću drugih lipida (fosfolipida u žuči i masnim kiselinama u crijevu), koji smanjuju monometrijsku koncentraciju žučnih kiselina, i drugo, uz prisustvo glikolipida i holesterola u apikalnim membranama epitelnih stanica, što povećava otpornost epitelnih stanica na oštećenje efekti anjona žučne kiseline.
Holne, cenodeoksiholne i deoksiholne kiseline apsorbuju se i prolaze kroz enterohepatičku cirkulaciju do 6-10 puta dnevno. Litoholna kiselina se slabo apsorbuje i njena količina u žuči je mala. Bila žučnih kiselina je normalno oko 2,5 g, a dnevna proizvodnja primarnih žučnih kiselina, holnih i cenoodeksikolnih, je u prosjeku oko 330 i 280 mg, redom.
Regulacija formiranja žuči
Kao što je poznato, formiranje žuči se odvija kontinuirano, ali intenzitet ovog procesa varira. Povećajte unos hrane u žuči i prihvatili hranu. Refleksni efekat na kolerezu javlja se tokom stimulacije receptora gastrointestinalnog trakta (GIT), unutrašnjih organa i uslovljenih refleksnih efekata.
Vegetativna regulacija je obezbeđena parasimpatičkom holinergikom (povećanje žučne formacije) i simpatičnim adrenergičkim nervnim vlaknima (smanjuju formiranje žuči).
Humoralnu regulaciju provodi sama žuča zbog prisutnosti enterohepatičke cirkulacije žučnih kiselina i mehanizma negativne povratne sprege (vidi gore). Secretin stimuliše sekreciju žuči, sekreciju vode i elektrolita u svom sastavu. Glukagon, gastrin i kolecistokinin imaju slabiji stimulativni efekat.
Izlučivanje bilijara
Izlučivanje bilijara smatra se neobičnim načinom uklanjanja molekula i jona iz tijela koji se ne mogu izlučivati kroz bubrege. Najznačajniji među njima su holesterol (kao takav i u obliku žučnih kiselina) i bilirubina, kao i joni bakra, gvožđa itd.
Glavne komponente izlučivanja žuči
Kolesterol, praktično nerastvorljiv u vodi, transportuje se u sastavu mešanih micela formiranih, kao što je gore navedeno, žučnim kiselinama, fosfolipidima i samim holesterolom.
Molekuli fosfolipida obavljaju dvije važne funkcije. Prvo, oni značajno povećavaju micelarnu rastvorljivost holesterola, jer mešane micele koje sadrže fosfolipide rastvaraju mnogo više holesterola od jednostavnih micela koje sadrže samo molekule žučne kiseline. Drugo, prisustvo fosfolipida u žuči smanjuje kritičnu koncentraciju micelizacije i monometrijsku koncentraciju žučnih kiselina. Kao rezultat toga, smanjena je površinska aktivnost i citotoksičnost jetrene žuči.
Sa protokom žuči, mešane micele ulaze u tanko crevo, gde je dalja transformacija njihovih sastavnih komponenti različita. Žučne kiseline rastvaraju lipide, osiguravajući njihovu apsorpciju i apsorbuju se u udaljenijim dijelovima crijeva. Žučni fosfolipidi, nerastvorljivi u vodi, hidrolizuju se u crevima i nisu uključeni u enterohepatičku cirkulaciju. Žučne kiseline regulišu izlučivanje i stimulišu sintezu.
Pošto se apsorbuje približno 1/3 holesterola, 2/3 se izlučuje. Kod odraslih osoba, ravnoteža holesterola se osigurava njegovim oslobađanjem (oko 600 mg / dan) ili žučnim kiselinama (oko 400 mg / dan). Istovremeno, enterohepatična cirkulacija žučnih kiselina se smatra odloženim načinom izlučivanja holesterola.
Treba napomenuti da u poređenju sa životinjama, muškarac ima veći udeo holesterola u žuči. To se smatra rezultatom neispravne konverzije holesterola u žučne kiseline, kao i relativno niskom stopom sekrecije žučnih kiselina. Dakle, žuči su prezasićeni holesterolom u oko 25% starijih osoba, au 10-15% populacije nastaju kolesterola (holelitijaza).
Kao što je gore navedeno, sinteza žučnih kiselina iz holesterola reguliše se mehanizmom negativne povratne sprege.
Žuči su glavni način uklanjanja bilirubina iz organizma. Bilirubin - Konačni proizvod razgradnje hema, u njegovoj hemijskoj strukturi je tetraprol. Najveća količina bilirubina (80-85%) dolazi od raspadajućeg hemoglobina starenja crvenih krvnih zrnaca i preuranjenog raspadanja novoformiranih crvenih krvnih zrnaca u koštanoj srži ili cirkulacijskom sloju (takozvana neefikasna eritropoeza). Ostatak bilirubina nastaje kao rezultat razaranja drugih proteina koji sadrže hem (npr. Citokrom P-450, itd.) U jetri i mnogo manje u ekstrahepatičnom tkivu. Treba napomenuti da se katabolizam eritrocitnog hemoglobina u bilirubin primarno javlja u makrofagima slezine, jetre i koštane srži.
Izvori formiranja bilirubina (u procentima)

Kao što je poznato, nekonjugirani bilirubin je hidrofoban (nerastvorljiv u vodi) i potencijalno toksična supstanca koja cirkuliše u plazmi u stanju blisko vezanom za albumin i ne može se izlučiti urinom. Sposobnost organizma da ukloni bilirubin je povezana sa uklanjanjem potonjeg iz krvne plazme u ćeliji jetre, a zatim konjugacijom sa glukuronskom kiselinom i oslobađanjem u žuči već rastvorljivog u vodi (vezani ili direktni bilirubin). Proces konjugacije odvija se preko mikrosomalnog enzima uridin difosfat glukuronil transferaze. Konjugacija bilirubina sa glukuronskom kiselinom ima najvažniji fiziološki značaj, međutim, konjugacija bilirubina sa sulfatima, glukozom i ksilozom javlja se u malim količinama.
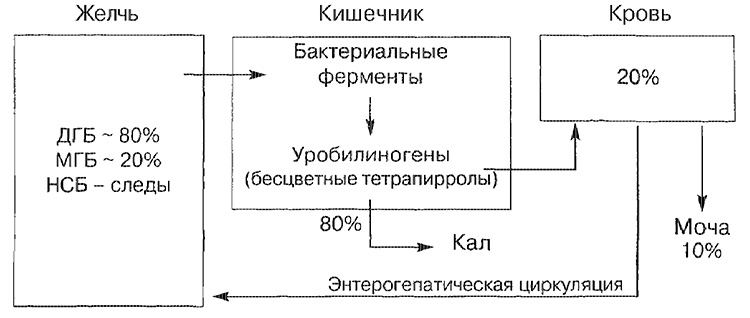
Glavni transportni putevi za bilirubin
NSB - nevezani bilirubin; MGB - bilirubin monoglukuronid; DGB - diglukuronid bilirubin.
Izlučivanje konjugiranog bilirubina u žučne tubule odvija se uz učešće porodice ATP-ovisnih multispecifičnih transportnih proteina za organske anione.
Većina bilirubin bilirubina (oko 80%) je zastupljena u obliku diglukuronid bilirubina, manji dio - u obliku monoglukuronida, a samo mali dio je predstavljen nevezanom formom. Bilirubin, koji je ušao u crijevo, razgrađuje se u terminalnom dijelu tankog crijeva i u debelom crijevu bakterijskim enzimima (beta-glukuronidaza), koji se pretvara u bezbojni tetapirol (urobilinogen). Oko 20% formiranih urobilinogena se resorbira i nužno se izlučuje u urinu, kao iu žuči (enterohepatički krug cirkulacije).
Većina autora se slaže da normalna koncentracija ukupnog serumskog bilirubina, određena Van Der Bergh diazo reakcijom, obično ne prelazi 1 mg% (0.3-1 mg%, ili 5-17 µmol / l). Samo manje od 5% bilirubina je prikazano u vezanom obliku. Povećanje nivoa bilirubina u krvi (hiperbilirubinemija) i njegova akumulacija u tkivima dovodi do pojave žutice, koja po pravilu postaje vidljiva na vrijednostima koje prelaze 2,5-3 mg%.
Kao što je gore navedeno, bilirubin cirkuliše u krvi u stanju vezanom za proteine i jedva prodire u tkivne fluide sa niskim sadržajem proteina. U tom smislu, eksudati su više žute boje nego transudati. Bilirubin se dobro veže za elastično tkivo, što objašnjava rano žuto bojenje sklera, kožnih i vaskularnih zidova kod hiperbilirubinemije. Produžena holestaza dovodi do pojave zelenkaste boje kože, što se objašnjava taloženjem biliverdina.
Prikladno je naglasiti da pritisak u žučnim kanalima, na čijoj se pozadini javlja sekrecija žuči, normalno iznosi 150-200 mm vode. Art. Povećava se na 350 mm vode. Art. inhibira sekreciju žuči, što dovodi do razvoja žutice. U slučaju potpunog prestanka izlučivanja bilirubina i žučnih kiselina, žuč postaje bezbojna (tzv. Bijela žuč).
Osim toga, žuč je način izlučivanja biljne masti, lipofilni lekovi i njihovi metaboliti, razne ksenobiotike koji su prisutni u biljkama, lipofilni metaboliti, vitamini rastvorljivi u mastima i steroidni hormoni.
Ravnoteža željeza i bakra telo je podržano i zbog žučne ekskrecije ovih metala. Oba kationa se luče u žuč pomoću ATP-stimulisane kanalikularne pumpe. Sa žuči se javlja i oslobađanje drugih metala.
Izlučivanje žuči
Struja žuči u bilijarnom sistemu je uzrokovana razlikama u pritiscima u različitim sekcijama i duodenumu, sfinkteru, kontrakciji glatkih mišićnih vlakana crvotočine i kanalima (prva pojedinačna mišićna vlakna pojavljuju se u intrahepatičnim žučnim kanalima koji se nalaze u neposrednoj blizini vrata jetre). Aktivnost čitavog sistema je normalno dobro koordinirana i pod kontrolom nervnog i humoralnog mehanizma.
Kod žučnih kamenaca koncentracije žuči se talože i povećavaju sa 10-strukim povećanjem koncentracije lipida, žučnih soli, žučnih pigmenata, itd. U pravom trenutku, oslobađaju se u OZP i dalje u duodenum. Prolaz žučnih koordinata sfinkter aparata. Izvan probavnog procesa u dvanaesniku, sfinkter bočice je zatvoren. U ovom trenutku, RI i cistiĉni kanal su opušteni, što omogućava da žuĉa koja se proizvodi u jetri ulazi u lumen RI. Sluznica ZH upija vodu, ione. Istovremeno, žuč postaje koncentrisana. Proizvodi sluzi omogućavaju da žuč bude u koloidnom stanju.
Pritisak u lumenu žlijezde u mirovanju mnogo je manji nego u žučnim kanalima i iznosi 60-185 mm vode. Art. Razlika u pritisku je fiziološka osnova protoka žuči u žučnoj kesici sa zatvorenim Oddi sphincterom. U procesu varenja uslijed smanjenja GF tlaka povećava se na 150-260 mm vode. st, osiguravajući protok žuči u duodenum kroz opuštene ampule sfinktera. Kada žuč počne da teče u duodenum, pritisak u kanalima se postepeno smanjuje (sa svakim obrokom sadržaj masti se smanjuje 1-2 puta).
Period primarne reakcije bilijarnog sistema, zbog vrste, mirisa hrane i njenog prijema, traje oko 7-10 minuta. Zatim dolazi period evakuacije (glavni ili period pražnjenja ZHP), tokom kojeg, na pozadini izmjene kontrakcija i relaksacije ZP, žučna kesica, a zatim jetrena žuca izlaze u duodenum.
Refleksna stimulacija holekineze (uslovno i bezuslovno-refleksno) nastaje zbog receptora koji se nalaze u usnoj šupljini, želucu i dvanaestopalačnom crevu, kroz vagusne živce. Humoralna regulacija se uglavnom provodi pod utjecajem kolecistokinina, koji ima stimulirajući učinak na GF, uzrokujući njegovo smanjenje. Pored toga, slabi rezovi u GI javljaju se pod uticajem gastrina, sekretina i Bombezina. Naprotiv, glukagon, kalcitonin, antiholecistokinin, vasointestinalni peptid i pankreasni polipeptid inhibiraju smanjenje GI.
Žumanjci, masti, mlijeko i meso su moćni stimulansi izlučivanja žuči.
Poremećaji sfinkter aparata i gonadalni motilitet mogu uzrokovati promjenu smjera i brzine žučnog toka, diskinezije, refluksa izlučivanja pankreasa i sadržaja duodenuma u ACP, te žuči u kanalu pankreasa, što je osnova za razvoj brojnih patoloških stanja i bolesti.
Žučne soli, konjugirani bilirubin, holesterol, fosfolipidi, proteini, elektroliti i voda se luče hepatocitima u žučnim kanalićima. Aparat za sekreciju žuči uključuje transportne proteine tubularna membrana, unutarćelijske organelei strukture citoskelet. Uske kontakteizmeđu hepatocita, lumen tubula je odvojen od cirkulacijskog sistema jetre.
Cevasta membrana sadrži transportne proteine za žučne kiseline, bilirubin, katione i anione. Microvilli povećava njegovu površinu. Organele su predstavljeni Golgijevim aparatom i lizosomima. Uz pomoć vezikula, proteini (npr. IgA) se prenose iz sinusoidalne membrane do kanalikularne membrane, transport transportnih proteina sintetiziranih u ćeliji za holesterol, fosfolipide i, moguće, žučne kiseline iz mikrosoma do tubularne membrane.
Citoplazma hepatocita oko tubula sadrži strukturu citoskeleta: mikrotubule, mikrofilamentii srednji filamenti.
Mikrotubule se formiraju polimerizacijom tubulina i formiraju mrežu unutar ćelije, posebno u blizini bazolateralne membrane i Golgijevog aparata, sudjelujući u vezikularnom transportu posredovanom receptorom, sekrecijom lipida i pod određenim uvjetima - žučnim kiselinama. Stvaranje mikrotubula inhibirano je kolhicinom.
U konstrukciji mikrofilamena učestvuju interakcijski polimerizovani (F) i slobodni (G) aktini. Mikrofilamenti, koncentrirani oko tubularne membrane, određuju kontraktilnost i motilitet tubula. Phalloidin, koji poboljšava polimerizaciju aktina, i citokalasin B, koji ga slabi, inhibira pokretljivost tubula i izaziva holestazu.
Intermedijarni filamenti se sastoje od citokeratina i formiraju mrežu između membrana plazme, nukleusa, intracelularnih organela i drugih struktura citoskeleta. Pucanje intermedijernih filamenata dovodi do prekida intracelularnih transportnih procesa i uništenja lumena tubula.
Voda i elektroliti utječu na sastav tubularne sekrecije, prodirući kroz uske kontakte između hepatocita zbog osmotskog gradijenta između cjevastog lumena i disesnih prostora. (paracelularna struja).Integritet tesnih kontakata zavisi od prisustva na unutrašnjoj površini plazma membrane proteina ZO-1 sa molekulskom masom od 225 kDa. Puknuće tesnih kontakata prati ulazak rastvorenih većih molekula u kanalikule, što dovodi do gubitka osmotskog gradijenta i razvoja kolestaze. Istovremeno se može uočiti regurgitacija kanalikularne žuči u sinusoidi.
Žučni kanalići ulaze u duktule, ponekad se nazivaju holangioli ili Goeringovi kanali. Duktule se nalaze uglavnom u portalnim područjima i ulivaju se u interlobularne žučne kanale, koji su prvi u bilijarnim traktima praćeni granama jetrene arterije i portalne vene i nalaze se u sastavu portalnih trijada. Interlobularni kanali, koji se spajaju, formiraju septalne kanale dok se ne pojave dva glavna jetrena kanala, koji izlaze iz desnog i lijevog režnja u portalnoj fisuri jetre.
Sekcija žuči
Nastajanje žuči nastaje uz učešće niza nestabilnih transportnih procesa. Njegova sekrecija je relativno nezavisna od perfuzijskog pritiska. Ukupna struja žuči kod ljudi je oko 600 ml / dan. Hepatociti osiguravaju izlučivanje dvije frakcije žuči: ovise o žučnim kiselinama ("225 ml / dan) i ne ovise o njima (" 225 ml / dan). Preostalih 150 ml / dan izlučuju ćelije žučnog kanala.
Izlučivanje soli žučnih kiselina je najvažniji faktor u formiranju žuči (frakcija zavisi od žučnih kiselina).Voda se kreće u svjetlu osmotski aktivnih soli žučnih kiselina. Promene osmotske aktivnosti mogu regulisati protok vode u žuč. Postoji jasna korelacija između izlučivanja soli žučnih kiselina i protoka žuči.
Postojanje žučne frakcije, koja ne zavisi od žučnih kiselina, dokazana je mogućnošću formiranja žuči koja ne sadrži soli žučnih kiselina. Prema tome, moguće je nastaviti protok žuči, uprkos odsustvu izlučivanja soli žučnih kiselina; lučenje vode u ovom slučaju je zbog drugih osmotski aktivnih rastvornih supstanci, kao što su glutation i bikarbonati.
Ćelijski mehanizmi sekrecije žuči
Hepatocit je polarna sekretorna epitelna ćelija koja ima bazolateralne (sinusoidne i lateralne) i apikalne (tubularne) membrane.
Formiranje žuči uključuje hvatanje žučnih kiselina i drugih organskih i neorganskih jona, njihov transport kroz bazolateralnu (sinusoidnu) membranu, citoplazmu i tubularnu membranu. Ovaj proces je praćen osmotskom filtracijom vode sadržane u hepatocitnom i paracelularnom prostoru. Identifikacija i karakterizacija transportnih proteina sinusoidnih i tubularnih membrana je kompleksna. Posebno je teško proučavanje sekretornog aparata tubula, međutim, do sada je razvijena tehnika za dobijanje duplih hepatocita u kratkotrajnoj kulturi i dokazana je pouzdanost u mnogim istraživanjima.Kloniranje transportnih proteina omogućava nam da karakteriziramo funkciju svakog od njih posebno.
Proces formiranja žuči zavisi od „prisustva određenih proteina nosača u bazolateralnim i tubularnim membranama. Na +, K igra ulogu pokretačke sile sekrecije. + - ATPaza bazolateralne membrane, obezbeđujući hemijski gradijent i potencijalnu razliku između hepatocita i okolnog prostora. Na +, K + - ATPaza razmenjuje tri intracelularna natrijumova jona za dva ekstracelularna kalijumova jona, održavajući gradijent koncentracije natrijuma (visoko spolja, nisko unutra) i kalijuma (nisko spolja, visoko iznutra). Kao rezultat, sadržaj ćelija ima negativan naboj (–35 mV) u poređenju sa ekstracelularnim prostorom, što olakšava hvatanje pozitivno naelektrisanih iona i izlučivanje negativno nabijenih iona. Na +, K + -ATPaza se ne otkriva u tubularnoj membrani. Protok membrane može da utiče na aktivnost enzima.
Snimite na površinu sinusoidalne membrane
Bazolateralna (sinusoidna) membrana ima mnogo transportnih sistema za hvatanje organskih aniona, čija se supstratna specifičnost delimično preklapa. Karakterizacija proteina nosača prethodno je data na osnovu proučavanja ćelija životinja. Nedavno kloniranje humanih transportnih proteina omogućilo je bolje opisivanje njihove funkcije. Transportni protein za organske anione (organski protein koji transportuje anion - OATP) je nezavisan od natrijuma, nosi molekule brojnih jedinjenja, uključujući žučne kiseline, bromsulfalein i, verovatno, bilirubin. Smatra se da transport bilirubina u hepatocite obavljaju i drugi nosioci. Hvatanje žučnih kiselina konjugiranih s taurinom (ili glicinom) vrši se natrijevim / tauroholatnim transportnim proteinom (natrijev / žučna kiselina kotransportni protein - NTCP).
Prenos jona kroz bazolateralnu membranu uključuje protein koji razmjenjuje Na + / H + i regulira pH unutar stanice. Ovu funkciju obavlja i cotransport protein za Na + / HCO 3 -. Na površini bazolateralne membrane, uhvaćeni su i sulfati, neesterificirane masne kiseline i organski kationi.
Intracelularni transport
Transport žučnih kiselina u hepatocitima vrši se pomoću citosolnih proteina, među kojima glavnu ulogu ima Z-hidroksisteroid dehidrogenaza. Glutation-S-transferaza i proteini koji vezuju masne kiseline su manje važni. Endoplazmatski retikulum i Golgijev aparat su uključeni u prijenos žučne kiseline. Očigledno, vezikularni transport se aktivira samo kada se žučne kiseline značajno ubrizgavaju u ćeliju (u koncentracijama koje prelaze fiziološke).
Transport proteina u tečnoj fazi i ligandima, kao što su IgA i lipoproteini niske gustine, vrši se vezikularnom transcitozom. Vreme prenosa od bazolateralne do tubularne membrane je oko 10 minuta. Ovaj mehanizam je odgovoran samo za mali dio ukupne struje žuči i ovisi o stanju mikrotubula.
Tubularna sekrecija
Cevasta membrana je specijalizovana sekcija plazma membrane hepatocita koja sadrži transportne proteine (uglavnom ATP-zavisne) odgovorne za prenos molekula u žuč protiv gradijenta koncentracije. Enzimi kao što su alkalna fosfataza, GGTP su takođe lokalizovani u kanaličnoj membrani. Prenos glukuronida i glutation-S-konjugata (na primer, bilirubin diglukuronida) vrši se uz pomoć kanalikularnog multispecifičnog transportnog proteina za organske anione (cMOAT), transporta žučnih kiselina sa kanalikularnim transportnim proteinom za žučne kiseline (kanalikularni b) transporter - SWAT), čija je funkcija djelomično kontrolirana negativnim unutarstaničnim potencijalom. Struja žuči, koja ne zavisi od žučnih kiselina, očigledno je određena transportom glutaiona, kao i kanaličnom sekrecijom bikarbonata, moguće uz učešće proteina koji razmjenjuju Cl - / HCO 3 -.
Važnu ulogu u transportu supstanci kroz tubularnu membranu ima dva enzima porodice P-glikoproteina; oba enzima ovise o ATP. Protein 1 za višestruku otpornost na više lijekova (MDR1) protein 1 za rezistenciju na više lijekova prenosi organske katione i također uklanja citostatike iz stanica raka, uzrokujući njihovu otpornost na kemoterapiju (otuda i ime proteina). Endogeni supstrat MDR1 je nepoznat. MDR3 prenosi fosfolipide i djeluje kao flippaza za fosfatidilkolin. Funkcija MDR3 i njena važnost za sekreciju fosfolipida u žuči su prerađeni u eksperimentima na miševima kojima nedostaje mdr2-P-glikoprotein (analogan humanom MDR3). U odsustvu fosfolipida u žuči, žučne kiseline uzrokuju oštećenje bilijarnog epitela, upalu duktula i periduktularnu fibrozu.
Voda i neorganski joni (posebno natrijum) izlučuju se u bilijarne kapilare duž osmotskog gradijenta difuzijom kroz negativno nabijene polupropustljive, čvrste kontakte.
Sekcija žuči se reguliše mnogim hormonima i sekundarnim glasnicima, uključujući cAMP i protein kinazu C. Povećanje koncentracije intracelularnog kalcijuma inhibira sekreciju žuči. Prolaz žuči kroz tubule je zbog mikrofilamenata koji obezbeđuju pokretljivost i kontrakciju tubula.
Duktilni sekret
Epitelne ćelije distalnih kanala proizvode tajnu koja je bogata bikarbonatom i koja mijenja sastav tubularne žuči (tzv. duktularna struja, žuč).Proces sekrecije proizvodi cAMP, neke proteinske membrane, uključujući protein koji razmenjuje Cl - / HCO 3 -, i regulator transmembranske provodljivosti kod cistične fibroze -membranski kanal za Cl -, podesivi cAMP. Duktularna sekrecija se stimuliše sekretinom.
Pretpostavlja se da urtodeoksiholnu kiselinu aktivno apsorbuju duktularne ćelije, razmjenjuju se bikarbonati, recikliraju u jetri i zatim izlučuju ponovo u žuč ("holhepatski šant"). Možda ovo objašnjava choleretic efekat ursodeoksiholne kiseline, praćen visokom sekrecijom bilijarnog bikarbonata u eksperimentalnoj cirozi.
Pritisak u žučnim kanalima, kod kojeg dolazi do izlučivanja žuči, obično je 15-25 cm vode. Art. Povećajte pritisak na 35 cm vode. Art. dovodi do supresije sekrecije žuči, razvoja žutice. Lekcija bilirubina i žučnih kiselina može se potpuno zaustaviti, dok žuč postaje bezbojna (bijela žuč)i podseća na sluzokožu.
Žuči je izomotična plazma tekućina koja se sastoji od vode, elektrolita i organskih supstanci (žučnih kiselina, fosfolipida, holesterola, bilirubina). Žučne kiseline (ili njihove soli) su glavna organska komponenta žuči. Žučne kiseline ulaze u žuč iz dva izvora: (1) primarne žučne kiseline (holne i ceneodoksiholne), koje se sintetišu iz holesterola u jetri; (2) sekundarne žučne kiseline (deoksikolne, litoholne i ursodeoksiholne) nastaju djelovanjem crijevnih bakterija iz primarnih žučnih kiselina. Žučne kiseline se sastoje od dve važne komponente koje određuju njihove fiziološke i fizičko-hemijske osobine: (1) steroidno jezgro sa hidroksilnim supstituentima; (2) alifatski bočni lanac (slika 7-4).
Sl. 7-4.
Žučne kiseline se sastoje od dvije komponente - jezgre s hidroksilnim krajevima i alifatskog bočnog lanca. Na slici, holinska kiselina je prikazana kao primer tri-hidroksi kiseline (3a-, 7p-, 12p-OH). Drugi primjeri su žučne kiseline koje sadrže deoksiholat (3a-12, -OH), cenoodoksiholat (3a-7, -OH) i litoholat (3p-OH)
Kod većine sisara, primarne žučne kiseline sadrže od tri do sedam hidroksilnih supstituenata, čiji broj utiče na rastvorljivost u vodi (hidrofilnost). Ubrzo nakon formiranja, primarne žučne kiseline prolaze modifikaciju u terminalnoj karboksilnoj grupi. Ovo se dešava tokom jetrene faze enterohepatičke cirkulacije sekundarnih žučnih kiselina i njihove konjugacije sa glicinom ili taurinom. Prisustvo hidrofilnih (hidroksilnih komponenti i amidnih veza alifatskog bočnog lanca) i hidrofobnih (steroidnih jezgara) komponenti omogućava da konjugirani molekuli žučne kiseline djeluju kao amfoterni spoj. To im daje mogućnost da formiraju micele (polimolekularne agregate) iznad kritične koncentracije micelara. S druge strane, molekuli žučne kiseline su u stanju da rastvore druge amfoterne supstance (holesterol, fosfolipide) sa formiranjem mešanih micela. Ova uloga žučnih kiselina u obliku deterdženta je važna za stabilizaciju fizičko-hemijskog stanja žuči, varenja i apsorpcije masti.
Sinteza žučnih kiselina iz holesterola reguliše se negativnim mehanizmom povratne sprege, iako priroda regulacije na molekularnom i biokemijskom nivou još nije u potpunosti shvaćena. Mikrosomska 7-Hidroksilacija holesterola je kritični korak u sintezi žučnih kiselina. Cenodesoksiholna kiselina, koja se koristi za rastvaranje kamenaca žučne kese, inhibira sintezu žučnih kiselina i time povećava nivo holesterola u krvi. Kada se koristi ursodeoksiholna kiselina, takve promene se ne primećuju ni sa dugotrajnim tretmanom.
Formiranje žuči odvija se i na sinusoidalnoj i na tubularnoj površini membrane hepatocita i predstavlja intracelularni i paracelularni proces. Za razliku od glomerularne filtracije u bubrezima, koja pasivno pod dejstvom hidrostatskih sila, tokom formiranja žuči, dolazi do aktivnog transfera organskih i neorganskih komponenti u lumen tubula i pasivnog transporta vode. Dakle, procesi sekrecije žuči su slični procesima sekrecije u acinium pankreasa, epitelom bubrežnih tubula. Formiranje tubularne žuči se može podeliti na dva tipa (Slika 7-5): (1) formiranje žuči, u zavisnosti od sekrecije žučnih kiselina, definisane kao odnos količine žuči koja se izlučuje u tubule i količine izlučenih žučnih soli; (2) stvaranje žuči, nezavisno od izlučivanja žučne kiseline, koje se može predstaviti kao aktivno izlučivanje neorganskih elektrolita i drugih supstanci i koje se odražava na grafu kao y-presek ove linije. Drugim riječima, stvaranje žuči povezano sa lučenjem kiseline je brzina protoka žuči, ovisno o prisutnosti osmotski aktivnih žučnih soli u žučnim kanalima, i stvaranje žuči nepovezanih s izlučivanjem kiseline u odsustvu žučnih soli. Odnos brzine formiranja žuči i formiranja soli žučnih kiselina je nelinearan sa malim količinama izlučene žuči i ne može odgovarati linearnom odnosu prikazanom na slici. 7-5. Stoga, oba tipa žučne formacije treba smatrati međusobno povezanim pokazateljima formiranja žuči.
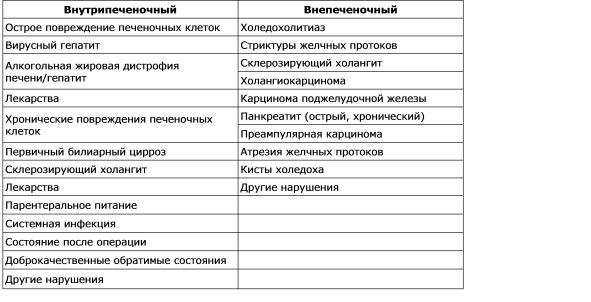
Kršenje formiranja žuči naziva se holestaza. Redoslijed patoloških, fizioloških i kliničkih manifestacija holestaze zavisi od uzroka. Morfološka studija materijala hepatobiopsije žuči otkrivena je u tubulima pericentralnih hepatocita, uočena je dilatacija tubula, a studija ultrastrukture otkriva smanjenje broja mikrovila. Holestaza se može definisati kao funkcionalni defekt u formiranju žuči na nivou hepatocita (intrahepatična holestaza), kao i organska ili mehanička narušena sekrecija i odliv žuči (extrahepatic cholestasis). Najčešći uzroci intrahepatične i ekstrahepatične holestaze su dati u tabeli. 7-2. Postoji nekoliko mehanizama koji igraju važnu ulogu u patogenezi intrahepatične holestaze: oštećenje i oštećenje funkcije sinusoidalne membrane; poremećaj funkcija intracelularnih organela u hepatocitima; oštećenje i ometanje tubularne membrane. Dakle, ne postoji jedinstveni mehanizam za holestazu u različitim kliničkim situacijama, a mnoštvo mehanizama može dovesti do različitih poremećaja. Klinički, holestaza se karakteriše povećanjem nivoa krvi mnogih supstanci, uključujući bilirubin, žučne soli, holesterol, koje se normalno izlučuju u žuč. Biohemijska analiza krvi sa holestazom dovodi do nesrazmernog povećanja aktivnosti alkalne fosfataze i koncentracije bilirubina paralelno sa promenama u nivou aminotransferaza, o čemu će biti reči u nastavku.
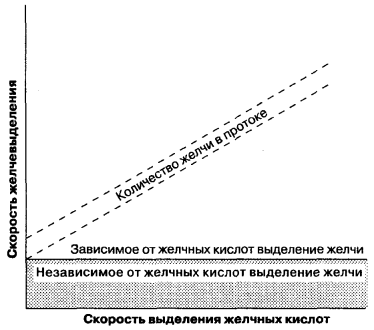
Sl. 7-5.
Nastanak žuči povezan sa sekrecijom žučne kiseline, formiranjem žuči nezavisno od izlučivanja žučne kiseline. (Autor: Moseley R.H., sekrecija žuči. U: Yamada T., Alpers D.H., Owyang C., Powell D.W., Silverstein F.E., eds. Textbook of Gastroenterology, 2. izdanje, Philadelphia: J. B. Lippincott, 1995: 387.)
Tabela 7 - 2.
U jetri se formira najvažniji probavni sok - žuč.
Žuči se proizvode hepatocitima aktivnim i pasivnim transportom vode, holesterola, bilirubina, kationa u njih. U hepatocitima iz holesterola formiraju se primarne žučne kiseline - holne i deoksikolične. Kompleks koji je rastvorljiv u vodi sintetiše se iz bilirubina i glukuronske kiseline. Oni ulaze u žučne kapilare i kanale, gde se žučne kiseline kombinuju sa glicinom i taurinom. Kao rezultat, nastaju glikolne i taurokolne kiseline. Natrijum bikarbonat se formira istim mehanizmima kao iu pankreasu.
Žuči se stalno proizvode u jetri. U svom danu se formira oko 1 litra. Hepatociti izlučuju primarnu ili hepatičnu žuč. Ova tečnost je zlatno-žuta alkalna reakcija. Njegov pH = 7,4 - 8,6. Sastoji se od 97,5% vode i 2,5% čvrstih materija. Suhi ostatak sadrži:
1. Mineralne supstance. Natrijum, kalijum, kalcijum kalcij, bikarbonat, fosfatni anioni, hlorni anioni.
2. Kisele kiseline - taurokolični i glikoholični.
3. Žučni pigmenti - bilirubin i njegov oksidovani oblik biliverdin. Bilirubin daje žučnu boju.
4. Holesterol i masne kiseline.
5. Urea, mokraćna kiselina, kreatinin.
Budući da je izvan probavnog sistema zatvoren sfinkter Oddi, koji se nalazi na ušću zajedničkog žučnog kanala, izlučena žuč se nakuplja u žučni mjehur. Ovdje se voda reabsorbira iz njega, a sadržaj osnovnih organskih komponenti i mucina se povećava za 5-10 puta. Dakle, cistična žuč sadrži 92% vode i 8% suvog ostatka. Tamnija je, deblja i viskoznija od jetre. Zbog ove koncentracije, mokraćna bešika može akumulirati žuč 12 sati. Tokom probave, sfinkter Oddi i sfinkter Lutkensa u vratu bešike su otvoreni. Bile ulazi u duodenum.
Bile Value:
1. Jabučne kiseline emulgiraju dio masti, pretvarajući velike čestice masti u fine kapljice.
2. Aktivira enzime intestinalnog i pankreasnog soka, posebno lipaze.
3. U kombinaciji sa žučnim kiselinama, apsorpcija dugolančanih masnih kiselina i vitamina topljivih u mastima nastaje kroz membranu enterocita.
4. Želčka promoviše resintezu triglicerida u enterocitima.
5.Inaktivira pepsine, a takođe neutralizira kiseli himem koji dolazi iz želuca. Time se osigurava prelazak sa probavnog trakta na želudac.
6.Stimulira izlučivanje pankreasnih i intestinalnih sokova, kao i proliferaciju i desquamation enterocita.
7. Jača intestinalni motilitet.
8. Ima bakteriostatski efekat na crevne mikroorganizme i na taj način sprečava razvoj gnojnih procesa u njemu.
Regulacija formiranja žuči i izlučivanja žuči se uglavnom vrši humoralnim mehanizmima, iako oni igraju određenu ulogu. Najsnažniji stimulator nastanka žuči u jetri je žučna kiselina, koja se apsorbira u krv iz crijeva. Takođe se pojačava sekretinom, što doprinosi povećanju natrijum bikarbonata u žuči. Vergusni nerv stimulira proizvodnju žuči, simpatički inhibira.
Kada himus uđe u duodenum, I-ćelije počinju da oslobađaju i-celije holcistokinin-pancreozimina. Posebno ovaj proces stimulišu masti, žumanjak i magnezijum sulfat. CCK-PZ jača kontrakcije glatkih mišića mokraćne bešike, žučnih puteva, ali opušta sfinktere Lutkensa i Oddija. Žuč se oslobađa u crijevo. Refleksni mehanizmi igraju malu ulogu. Chyme iritira kemoreceptore tankog crijeva. Impulsi iz njih ulaze u probavni centar medulle oblongata. Od njega su na vagusu u bilijarnom traktu. Sfinkteri se opuštaju i glatke mišiće mokraćnog mjehura se smanjuju. Promoviše izlučivanje žuči.
Najozbiljnije bolesti su hepatitis i ciroza. Najčešće je hepatitis rezultat infekcije (infektivni hepatitis A, B, C) i izlaganje toksičnim proizvodima (alkoholu). Kod hepatitisa zahvaćeni su hepatociti i oštećene su sve funkcije jetre. Ciroza je rezultat hepatitisa. Najčešća povreda bilijarne ekskrecije je holelitijaza. Glavninu žučnih kamenaca formira holesterol, jer je žuč takvih pacijenata prezasićena njima.
Jetra je najveći organ u telu i centralna je za metabolizam. Obavlja mnoge funkcije, učestvuje u metabolizmu proteina, ugljenih hidrata, masti, hormona i vitamina, kao iu neutralizaciji mnogih endogenih i egzogenih supstanci. Ovi procesi su opisani u udžbenicima fiziološke hemije, au ovom dijelu u vezi sa probavni sistem samo ćemo razmotriti izlučujuća funkcija jetretj. sekrecija žuči. Bile sastoji se od vode, mineralnih soli, sluzi, holesterola i lecitina i dva tipa specifičnih komponenti - žučnih kiselina i bilirubinskog pigmenta. Žučne kiseline su deterdženti, a njihova emulgirajuća aktivnost igra važnu ulogu u varenju lipida. Bilirubin je krajnji produkt razgradnje hemoglobina koji se eliminira iz organizma.
^
Bile formation
Funkcionalna anatomija.Ćelije jetre (hepatociti)oblikuju ploču u debljini jedne ćelije, odvojene uskim prorezima (Disse prostor)koji predstavlja završen
^ 766 DIO VIII. HRANA, PRESTANAK I EKSTRAKT
Po krvi sinusoidiekvivalentna kapilarama. U zidovima sinusoida postoje pore kroz koje prolaze tako velike makromolekule kao albumin i lipoproteini. U najmanjem tubuleŽučni kapilari omeđeni membranama plazme dvaju susjednih hepatocita - žuč se skuplja u veće goringovi kanalići,zidovi od kojih, kao i oni veći intercollege tubulei žučnih kanalaformirane kubičnim sekretornim ćelijama. Male tubule unutar jetrenih zdjelica i između njih se stapaju u veće, i na kraju se formiraju jetreni kanal.Iz ovog kanala odlazi cističnog kanalado žučne kese. Nakon spajanja, formiraju se jetreni i cistični kanali zajednički žučotvara se u duodenumu na vrhu papile Vatera, iza ili blizu kanala pankreasa (Slika 29.1).
^ Funkcije žuči.Bile obavlja mnoge važne funkcije. Sa njom prikazani su krajnji proizvodirazmenu, kao što je bilirubin, kao i lekove i toksine. Raspodjela sažuči holesteroligra važnu ulogu u regulaciji svoje ravnoteže. Žučne kiselineneophodna za emulzifikaciju i apsorpciju masti. Osim toga, žuč sadrži vodu, mineralne soli i sluz. Oko 24 sata dnevno 600 mlžuči, i 2/3 ove količine dolazi iz tubula, i 1/3 većih kanala.
^ Tubularna žuči formira se u približno jednakim količinama uz učešće dva različita mehanizma, zavisno od žuči i žuči (slika 29.27).
^ Izlučivanje koje zavisi od žučnih kiselina. Postoji bliska veza između protok žučii izlučivanje žučne kiseline.U kanalikularnoj žuči koncentracija žučnih kiselina je 100 puta veća nego u portalnoj krvi, pa se smatra da se izlučuju aktivni transport koji uključuje vektor.Prateći žučne kiseline duž osmotskog gradijenta, voda ulazi u tubule, tako da je žuč izotonična sa krvlju.
Postoje dva izvora žučnih kiselina. Prvo, sintetiziraju se de novo iz holesterola u hepatocitima uz učešće 7-hidroksilaze. Ovaj enzim igra ključnu ulogu i kontrolira brzinu sinteze žučnih kiselina putem mehanizma povratne sprege. Drugo, hepatociti mogu aktivno da apsorbuju žučne kiseline portalna krvi izlučuju ih u tubule (vidi takođe sliku 29.29). Ova ekstrakcija je veoma efikasna; sa jednim prolaskom krvi kroz jetru, 80% žučnih kiselina se izdvajaju iz nje. Zato
Koncentracija žučne kiseline u perifernoj krvi je mnogo niža nego u portalnom sistemu. Pošto se žučne kiseline iz krvi izvlače 6 puta brže nego što ulaze u tubule, to je drugi proces koji ograničava izlučivanje žučne kiseline.
^ Izolacija nezavisna od žučnih kiselina. U
Ovaj proces uključuje ione Na +, Cl -, HCO 3 - i vode. Pokretačka snaga je aktivni transport α + , moguće sa bikarbonatom. Izolacija koja je nezavisna od žučne kiseline stimuliše, naročito, secretin.
Pored žučne kiseline, aktivno se izlučuju bilirubin, holesterol i fosfolipidi (uglavnom lecitin) u kanalikule (Sl. 29.27). Nerastvorljiv u vodi ("Indirektni") bilirubin,većina se formira iz hemoglobina starih crvenih krvnih zrnaca, ulazi u hepatocite u obliku koloidnog agregata povezanog sa albuminom. Njegova dnevna formacija je oko 4 g / kg tjelesne težine, ili 200-300 mg / dan. U hepatocitima je 80% bilirubina konjugirano. saglukuronska kiselina i mala količina sumporne kiseline. U takvim konjugiranim
^ POGLAVLJE 29. FUNKCIJE GASTROINTESTINALNOG TRAKTA 767
Bilirubin se izlučuje u žuči. ("Direktni" bilirubin).Općenito, lijekovi i toksini se uklanjaju na isti način.
^ Modifikacija žuči u žučnim kanalima
(sl. 29.27). U kanalima gdje se kanali otvaraju, primarna žuči se podvrgavaju modifikaciji. Ovaj proces podseća na proces modifikacije glomerularnog filtrata u bubrežnim tubulima (str. 785), i na sličan način se izračunava. čišćenje,samo u slučaju žuči kao inertne supstance koristi se umjesto inulina eritritolili manitol,koje se luče u tubule, ali se ne resorbiraju. Slične studije su pokazale da se oko 180 ml žuči, odnosno 1/3 ukupne količine, izlučuje u kanale sa aktivnom sekrecijom NSO ^ ". secretin
^
Žuč u jetri i žuči
Sastav jetrene žuči(tab. 29.3). Žuč koja se izlučuje jetrom brzinom od 0,4 ml / min ima zlatnu boju, što se objašnjava prisustvom bilirubina u njemu. Koncentracija elektrolita u ovoj žuči je ista kao u plazmi, izuzev činjenice da sadrži dvostruko više HCO3 i nešto manje od CI. U isto vreme, sastav organskih supstanci žuči se veoma razlikuje od plazme, jer su u žuči \\ t zastupljene gotovo isključivo žučnim kiselinama, holesterolom i fosfolipidima.
 |
^ Žučne kiselinenastaju u jetri iz holesterola kao rezultat njegove hidroksilacije i dodavanja karboksilne grupe. Kiseline koje se formiraju u jetri su primarne žučne kiseline;to uključuje chenodeoxycholic(dioksim) i holevaya(trioxyforma) kiselina.U jetri nisu u slobodnom obliku, već u obliku konjugata sa glicinom i taurinom, a konjugati sa glicinom se formiraju tri puta više, jer je količina taurina ograničena. Konjugirane žučne kiseline su topivije u vodi,
Nevezani i imaju veću sposobnost disocijacije i formiranja žučnih soli sa kationima, uglavnom sa Na + jonima. U kiselom okruženju (pH 4.0), soli žučnih kiselina su nerastvorljive i talože se, ali kod fizioloških pH vrijednosti (u tankom crijevu), one su dobro topljive.
U distalnom delu ileuma i debelog creva, neke od soli primarnih žučnih kiselina prolaze dehidroksilaciju pod dejstvom anaerobnih bakterija i pretvaraju se u sekundarna žučna kiselina-litokol(monooxyform) i deoxycholic(dioxyform). Cenoodeksikolne, holne i deoksiholne kiseline prisutne su u omjeru 2: 2: 1. Litoholna kiselina je prisutna samo u nekim frakcijama, jer se većina izlučuje.
Emulgirajući učinak žučnih kiselina na masti uglavnom se zasniva na njihovoj sposobnosti da se formiraju micelles.Molekuli žučne kiseline imaju trodimenzionalnu strukturu u kojoj su hidrofilne karboksilne i hidroksilne grupe na istoj strani molekula, a hidrofobni dio molekule (steroidna jezgra, metilne grupe) je na suprotnoj strani, zbog čega molekule žučnih kiselina posjeduju i hidrofilnoi lipofilna svojstva.Zbog ove strukture, molekuli žučne kiseline djeluju kao deterdženti: na granici lipidne i vodene faze, oni formiraju gotovo monomolekularni film u kojem se hidrofilne grupe pretvaraju u vodenu, a lipofilnu u lipidnu fazu. U vodenoj fazi, žučne kiseline formiraju uređene agregate. -micelspod uslovom da njihova koncentracija dostigne određeni nivo, naziva se kritična koncentracija micela(1-2 mmol / l). Unutrašnja, lipofilna regija micela može sadržavati lipidina primer, holesterol i fosfolipidi; takve micele se nazivaju mješovite (slika 29.28). Sam holesterol je nerastvorljiv u vodi, ali može biti u rastvoru u sastavu micela. Ako njegova koncentracija prelazi kapacitet micela, on formira kristalni talog; ovaj proces je osnova formiranja kolesterola u žuči (str. 769).
^ Sastav cistične žuči (tab. 29.3). Kapacitet žučne kese je samo 50-60 ml. Istovremeno, jetra izlučuje žuč u količini od 600 ml / dan, a polovina te količine prolazi kroz žučnu bešiku prije ulaska u tanko crijevo. Razlika između količine žuči koja ulazi u žučnu kesicu i njenog kapaciteta se kompenzira visoko efikasna reapsorpcijavode u žučnoj kesi. U roku od nekoliko sati, 90% vode se može resorbovati iz žuči. Sa
^
768 DIO VIII. HRANA, PRESTANAK I EKSTRAKT
Ova organska materija ostaje u žučnoj kesici i povećava se koncentracija žuči. Pokretačka snaga reapsorpcije je aktivan transport N * ionauz učešće "pumpe" ugrađene u bazalne i lateralne membrane ćelija i aktivirane pomoću (Na + -K +) - ATPaze. Slijedeći Na + ioni, Cl - i HCO 3 - ioni difundiraju u smjeru električnog gradijenta ili se transportiraju nosačima. Kao rezultat reapsorpcije HCO 3 - pH žučnog mjehura se smanjuje na 6,5 naspram 8,2 u jetrenoj žuči. Kao rezultat stvaranja visoke koncentracije Na + iona u epitelu žučne kese u ekstracelularnom prostoru javlja se osmotski gradijent, koji dovodi do pumpanja vode, koja zatim ulazi u kapilare (str. 751).
^ Motilitet žučne kese. U stanju gladovanja, žuči se nakupljaju u žučnoj kesi i unutra
Vreme jede se dodeljuje kao rezultat kontrakcija žučne kese. Glavni stimulator kontraktilne aktivnosti žučne kese je cholecystokinin,izlučuje mukusna sluznica dvanaestopalačnog crijeva nakon prijema u njega himus koji sadrži masnoću. Kontrakcije žučne kese su takođe stimulisane u određenoj meri. lutajući nervi parasympatholytics.Počinju u roku od 2 minuta nakon što masna hrana dođe u kontakt sa crijevnom sluznicom, a nakon 15-90 minuta mjehur se potpuno isprazni. Motilitet žučne kese uključuje dva procesa. U početku se razvija tonička kontrakcija, uslijed čega se smanjuje promjer žučne kese, a zatim se na taj efekat primjenjuju periodične kontrakcije, čija je učestalost 2-6 / min. Kao rezultat ova dva procesa nastaje pritisak od 25–30 mm Hg. Art.
^
Enterohepatična cirkulacija
Cirkulacija žučne kiseline(sl. 29.29). Žučne kiseline se luče u duodenum kao mešane micele. Unatoč razrjeđivanju žučnih kiselina sadržajem želuca, njihova koncentracija u crijevu je oko 10 mmol / l i ostaje iznad kritične koncentracije formiranja micela. Ovde, pored holesterola i lecitina, micele uključuju produkte razlaganja hidrolitičke masti - masne kiselinei monogliceridi.Pri početnom kontaktu micela sa crevnim zidom
^
POGLAVLJE 29. FUNKCIJE GASTROINTESTINALNOG TRAKTA 769
Lipidi se šire kroz membranu četkaste granice u enterocite, a žučne kiseline ostaju u lumenu crijeva, ali uz daljnji prolaz kroz crijeva, žučne kiseline se apsorbiraju aktivnim i pasivnim transportom.
Oko 50% žučnih kiselina se apsorbira u crevima pasivno. Kao rezultat cijepanja konjugata žučne kiseline i dehidroksilacije potonjih djelovanjem crijevnih bakterija, povećava se njihova topljivost u lipidima i olakšava se pasivna difuzija.
^ Aktivno usisavanje žučne kiseline javljaju se isključivo u terminalnom ileumu - rijetka pojava, poznata samo za apsorpciju vitamina B12. Samo one žučne kiseline koje imaju visok polaritet koje ometaju njihovu pasivnu apsorpciju, kao što su konjugati taurina, podložne su aktivnoj apsorpciji. Proces apsorpcije žučnih kiselina u terminalnom ileumu karakterišu tipični znaci aktivnog transporta: kinetika zasićenja i kompetitivna inhibicija. Mala količina žučnih kiselina (7-20%) nije uključena ni u aktivnu ni u pasivnu apsorpciju i eliminisana je iz organizma.
Čini se da prisustvo žučnih kiselina u kolonu igra važnu ulogu u regulaciji konzistentnosti fekalija. Kada se koncentracija dioksidne kiseline u debelom crijevu iznad 3 mmol / l, značajna količina elektrolita i vode izlučuje u crijevni lumen, što dovodi do proljeva. Izraženi oblik ovoga "Hologennogo" dijarejamože se primijetiti kod resekcije ili bolesti terminalnog ileuma, te za njegovo liječenje upotrebom vezanja žučnih kiselina pomoću izmjenjivača iona holestiramina.
Kada se proguta apsorbuje žučne kiseline do jetrekonjugati su tek formirani, a neke sekundarne žučne kiseline se podvrgavaju hidroksilaciji. Gubitak žučnih kiselina u fecesu (0,2-0,6 g / dan) kompenzira se njihovom sintezom.
^ Ukupna količina žučnih kiselina u tijelu je oko 3,0 g. Ova količina nije dovoljna da osigura lipolizu nakon obroka; posebno, konzumiranje masne hrane zahtijeva 5 puta više žučnih kiselina. Međutim, telo nije oskudno sa žučnim kiselinama, jer cirkuliše kroz creva i jetru mnogo puta. (enterohepatična cirkulacija).Učestalost kojom bazena žučnih kiselina čini puni ciklus zavisi od režima ishrane i kreće se od 4 do 12 ciklusa dnevno.
^ Cirkulacija bilirubina. Gall pigment bilirubin,kao žučne kiseline i lipidi, ulazi u crevo u obliku glzhuronida. Samo mala količina ovog polarnog jedinjenja
Reapsorbira se u žučnoj kesi i tankom crijevu. U terminalnom ileumu i (uglavnom) u kolonu konjugat bilirubina se cijepa djelovanjem bakterijskih hidrolaza. Istovremeno se pretvara i bilirubin urobilinogenkoji, zajedno sa drugim produktima raspadanja bilirubina, daje smeđu boju izmetu. Manje od 20% urobilinogena se apsorbuje natrag, a od te količine, oko 90% ponovo ulazi u jetru i vraća se žuč, a preostalih 10% se izlučuje urinom.
Patofiziološki aspekti. Povećani nivo urobilinogena u urinu može ukazivati bolest jetre,praćeno kršenjem izlučivanja bilirubina. Potpuno odsustvo urobilinogena u mokraći, svetle boje fecesa i žutosti ukazuju potpuna blokada žučne kese;u ovoj situaciji, bilirubin uopšte ne ulazi u crijevo i ne stvara se urobilinogen.
Najpoznatija i široko rasprostranjena povreda normalne fiziologije bilijarnog sistema je taloženje holesterola sa formiranjem holesterola u žučnim kamencima. Holesterol, kao i lecitin, nalazi se u rastvorenom stanju samo kao deo mešanih micela. Ako koncentracija raste xo.jecmepo.ia takođe koncentracija žučnih kiselina ili lecitina postaje ispod kritičnih nivoa,precipitati holesterola. Među faktorima koji uzrokuju povećanje relativnog sadržaja holesterola su estrogeni, dijeta ugljikohidrata, prekomjerna težina i procesi koji smanjuju koncentraciju žučnih kiselina, kao što je upala ileuma. (Crohnova bolest)ili resekcija. U nekim slučajevima, oralna nadoknada žučne kiseline može biti dovoljna da se litogene žuči pretvore u alitogene, u kojima se kamenac holesterola može otopiti. Cenodeoksiholne i urodesoksiholne kiseline su najpogodnije za ovu svrhu, jer ne izazivaju dijareju.
Klinička manifestacija poremećaja metabolizma bilirubina je žutica. Žuta koža povezana je sa povišenim nivoima bilirubina u plazmi, koji se mogu pojaviti u sledećim slučajevima:
sa povećanim formiranjem bilirubina kao rezultat pojačanog sloma eritrocita (hemolitička žutica);
kao rezultat prekida procesa konjugacije ili transporta bilirubina u hepatocitima, kao, na primjer, žutica trudnaili gilbertova majčinska žutica;
sa odloženim odlivom žuči, na primer zbog žučnih kamenaca ili tumora lokalizovanih u predelu žučnog kanala (opstruktivna žutica).