The formation of bile. The physiology of bile formation. Gall formation
Bile formation occurs in the liver as a result of active and passive transport of substances (water, glucose, creatinine, electrolytes, hormones, vitamins, etc.) circulating in the blood, through cells and cell – cell contacts, as well as active secretion of bile components (bile acids) by hepatocytes and reabsorption of water and some substances from the small bile ducts and gallstones. The physiological significance of this process is diverse. Bile is primarily regarded as a digestive secret, since bile acids (mainly their organic anions) play a key role in the absorption of fats. Bile emulsifies fats, increasing the surface on which they are hydrolyzed under the influence of lipase. Under the action of bile is the dissolution of fat hydrolysis products. It promotes their absorption and resynthesis of triglycerides in enterocytes. Bile increases the activity of pancreatic and intestinal enzymes (especially lipases), enhances hydrolysis and absorption of proteins and carbohydrates.
In violation of the digestion of fats are badly digested and other food substances, because the fat envelops the smallest particles of food and prevents the action of enzymes on them. In such conditions, the activity of intestinal bacteria leads to increased processes of decay, fermentation and the formation of gas.
Regulatory effects are also characteristic of bile - stimulation of bile formation, biliary excretion, motor and secretory activity of the small intestine, as well as proliferation and desquamation of enterocytes. Bile inhibits the process of gastric digestion by neutralizing the acid that entered the duodenum and inactivating pepsin, preparing for digestion in the intestine. It also has a regulatory impact on the evacuation activity of the gastroduodenal complex. The role of bile in the absorption of fat-soluble vitamins (A, D, E and K), cholesterol, calcium salts from the intestinal lumen is important.
In addition, the formation and secretion of bile is considered as a peculiar way of excretion of certain molecules and ions that cannot be excreted through the kidneys. Among them, the most significant are cholesterol (displayed as free cholesterol, its esters and bile acids), bilirubin, as well as copper and iron molecules. Therefore, bile is considered as an excretory fluid.
Bile consists of 80% water and 20% of substances dissolved in it. The latter include bile acids and their salts (about 65%), biliary phospholipids (about 20%, mainly due to lecithin), proteins (about 5%), cholesterol (4%), conjugated bilirubin (0.3%), enzymes , immunoglobulins, as well as numerous exogenous and endogenous substances secreted with bile (plant styrenes, vitamins, hormones, drugs, toxins, metal ions - copper, iron, potassium, sodium, calcium, zinc, magnesium, mercury, etc.). On average, 600-700 ml of bile are secreted per day (from 250 to 1500 ml, approximately 10.5 ml per 1 kg of body weight). At the same time, about 500 ml / day of the total volume is provided by the secretion of hepatocytes and about 150 ml / day the cells of the bile ducts secrete.
Bile formation (choleresis) goes continuously, and the flow of bile into the duodenum (cholekinesis) happens periodically. On an empty stomach, bile enters the intestine in accordance with periodic hungry activity. During periods of rest, it goes to the WB, where it concentrates, somewhat changes its composition and is deposited. In addition to water and salts, cholesterol and free fatty acids are absorbed. In this regard, distinguish hepatic and gallbladder bile.
Bile has a small enzymatic activity; Hepatic bile pH is 7.3-8.0. Unlike intestinal contents, it contains almost no bacteria. The factors that ensure the sterility of bile include the presence of bile acids (bacteriostatic action), the rich content of immunoglobulins, the secretion of mucus, the relative poverty of bile energy substrates for bacteria.
Bile is a micellar solution. Cholesterol, practically insoluble in water, is transported in a dissolved state in the bile due to its micellar structure. This process is called colloidal dissolution - solubilization.
Bile acids are surface-active, amphipathic (both hydrophobic and hydrophilic) molecules capable of self-aggregation. In this case, thanks to a very narrow concentration rate, called the critical concentration of micellization, simple micelles are formed. Simple micelles have a pronounced ability to dissolve lipids, forming mixed micelles.
Mixed micelles are believed to have a cylindrical structure: the cylindrical stem is filled with polar lipids, and the bile acid molecules are located between the polar ends of lipid molecules with hydrophilic sides facing the water environment, which determines their hydrophilicity (water solubility).
Mixed micelles contain essential components - bile acids, located outside, phospholipids (mainly phosphatidylcholine - lecithin) and cholesterol, located inside the micelle.
The color of bile is yellowish-brown due to the presence of bilirubin, the largest part of which is presented in the form of diglucuronide bilirubin, the lesser - in the form of monoglucuronide bilirubin. Associated bilirubin is not part of the micelles.
Since the intercellular biliary tree is permeable to water, gallbladder and hepatic bile are isotonic.
Bile formation
Biliary secretion
Bile acids are the main component of biliary secretion, they are formed only in the liver. Primary bile acids - trihydroxycholic (cholium) and dihydroxycholic (chenodeoxycholic) are synthesized in cholesterol hepatocytes. Secondary bile acids (deoxycholic and in small quantities - lithocholic) are formed in the colon from primary as a result of bacterial modification of nuclear hydroxyl groups (7a-dehydroxylation under the control of anaerobic bacteria). Tertiary bile acids (mainly ursodeoxycholic) are formed in the liver by isomerization of secondary bile acids.
Synthesis of bile acids from cholesterol is regulated by a mechanism of negative feedback: a reduced return of bile acids to hepatocytes with portal blood leads to an increase in their biosynthesis; any increase in the synthesis of bile acids is accompanied by a corresponding increase in the formation of cholesterol.
Bile acids are conjugated in the liver with the amino acids glycine (about 80%) or taurine (about 20%). When carbohydrate-rich food is consumed, the amount of glycocholic acids increases in bile, and in high-protein diets, the amount of glycophenolic acids increases. Amino acid binding prevents the absorption of bile acids in the biliary tract and the initial sections of the small intestine (only in the terminal ileum and in the large intestine). Under the action of bacteria, hydrolysis of bile salts with the formation of bile acids, glycine or taurine is possible.
After the biosynthesis of bile acids, as noted above, their carboxyl group is bound to the amino group of glycine or taurine. From a chemical point of view, such a binding converts a weak acid to a stronger one. Bound bile acids are most soluble at low pH and more resistant to precipitation with calcium ions (Ca 2+) than unbound bile acids. From a biological point of view, the binding process makes bile acids incapable of penetration through cell membranes. Therefore, they are absorbed in the bile ducts or the small intestine, either in the presence of carrier molecules, or if the bile acids undergo bacterial cleavage. The only exception to this rule is glycine dihydroxy-associated bile acids, since they can be absorbed passively if they receive a hydrogen ion (H +).
Most of the bile acids (about 85-90%), entered with a current of bile into the small intestine, are absorbed into the blood. However, most of the associated bile acids that are secreted into the small intestine are absorbed intact. A smaller part of the acid is not absorbed intact, as it undergoes bacterial cleavage in the distal small intestine. They are absorbed passively and, returning to the liver, are again bound and secreted into the bile. The remaining 10-15% of bile acids are eliminated from the body mainly with feces. This loss of bile acids is compensated by their synthesis in hepatocytes.
This process of intestinal cleavage and reabsorption in the liver is a normal part of the metabolism of bile acids. A small portion of bile acids that have not been absorbed enters the colon. Here ends the splitting process. In addition, secondary bile acids are formed in the colon under the action of anaerobic bacteria (see above).
Deoxycholic and lithocholic acids are partially absorbed in the colon and re-enter the liver. After the return, the metabolism of these bile acids is different. Deoxycholic acid binds to glycine or taurine and circulates with the primary bile acids. It should be noted that in the elderly, deoxycholic acid is the main bile acid in the composition of bile. Litocholic acid not only binds to glycine or taurine, but additionally sulphates at the C-3 position. Such a “double” binding reduces the possibility of absorption in the intestine, as a result of which lithocholic acid is quickly lost from the composition of circulating bile acids - its content in the bile rarely exceeds 5%. Most (95%) bile acids, which are part of the bile of adults, are cholic, chenodesoxycholic and deoxycholic acids.
The process of binding bile acids in hepatocytes is very effective, so biliary bile acids are completely present in conjugated form. Subsequent bacterial cleavage and dehydroxylation lead to the fact that in the feces bile acids are in the unbound state.
The process of dehydroxylation of primary bile acids described above reduces their ability to dissolve in water. The formation of secondary bile acids in humans has practically no physiological significance, unlike animals. Excessive absorption of deoxycholic acid in the colon increases the risk of cholesterol gallstones. Litocholic acid is considered hepatotoxic. Animal experiments have shown that the accumulation of lithocholic acid in the liver leads to its defeat. However, it has not yet been proven that an increase in the absorption of this acid in humans leads to disruption of the liver.
Ursodeoxycholic acid, as well as deoxycholic acid, binds in the liver and circulates with the primary bile acids. However, the metabolic pathway of this acid is very short and the content of bound ursodeoxycholic acid in bile never exceeds 5% of the total amount of bile acids. It is believed that the formation of ursodeoxycholic acid does not have important physiological significance.
Bile acids are powerful solvents of fats, so they are cytotoxic in concentrations that reach the critical concentration of micellization. At the same time, bound chenodeoxycholic and deoxycholic acids present in bile are more toxic than those associated with cholic and ursodeoxycholic. Although bile acids are demonstratively cytotoxic in vitro, the epithelium of the bile ducts and small intestine is never damaged as a result of their high concentration. This is associated, firstly, with the presence of other lipids (phospholipids in the bile and fatty acids in the intestine), which reduce the monometric concentration of bile acids, and, secondly, with the presence of glycolipids and cholesterol in the apical membranes of epithelial cells, which increase the resistance of epithelial cells to the damaging the effects of bile acid anions.
Cholic, chenodeoxycholic and deoxycholic acids are absorbed and undergo enterohepatic circulation up to 6-10 times per day. Litocholic acid is poorly absorbed and its amount in the bile is small. The pool of bile acids is normally about 2.5 g, and the daily production of primary bile acids, cholic and chenodeoxycholic, is on average about 330 and 280 mg, respectively.
Regulation of bile formation
As is known, bile formation occurs continuously, but the intensity of this process varies. Enhance bile formation food intake and accepted food. A reflex effect on choleresis occurs during stimulation of the receptors in the gastrointestinal tract (GIT), internal organs, and conditioned reflex effects.
Vegetative regulation is provided by parasympathetic cholinergic (increase bile formation) and sympathetic adrenergic nerve fibers (reduce bile formation).
Humoral regulation is carried out by the bile itself due to the presence of enterohepatic circulation of bile acids and the mechanism of negative feedback (see above). Secretin stimulates the secretion of bile, the secretion of water and electrolytes in its composition. Glucagon, gastrin and cholecystokinin have a weaker stimulating effect.
Biliary excretion
Biliary excretion is considered as a peculiar way of removing molecules and ions from the body that cannot be excreted through the kidneys. The most important among them are cholesterol (as such and in the form of bile acids) and bilirubin, as well as ions of copper, iron, etc.
The main components of biliary excretion
Cholesterol, practically insoluble in water, is transported in the composition of mixed micelles, formed, as noted above, by bile acids, phospholipids and cholesterol itself.
Phospholipid molecules perform two important functions. First, they significantly increase the micellar solubility of cholesterol, since mixed micelles containing phospholipids solubilize much more cholesterol than simple micelles containing only bile acid molecules. Secondly, the presence of phospholipids in bile reduces the critical concentration of micellization and the monometric concentration of bile acids. As a result, the surface activity and cytotoxicity of hepatic bile are reduced.
With the flow of bile, mixed micelles enter the small intestine, where the further transformation of their constituent components is different. Bile acids dissolve lipids, ensuring their absorption, and are themselves absorbed in more distal parts of the intestine. Bile phospholipids, insoluble in water, hydrolyze in the intestine and are not involved in the enterohepatic circulation. Bile acids regulate their excretion and stimulate synthesis.
Since approximately 1/3 of cholesterol is absorbed, 2/3 of it is excreted. In an adult, the balance of cholesterol is ensured by its release as such (about 600 mg / day) or as bile acids (about 400 mg / day). At the same time, enterohepatic circulation of bile acids is considered as a delayed way of cholesterol excretion.
It should be noted that in comparison with animals, a higher proportion of cholesterol in bile is observed in humans. This is considered the result of defective conversion of cholesterol into bile acids, as well as the relatively low rate of secretion of bile acids. Therefore, bile is oversaturated with cholesterol in about 25% of the elderly, and in 10-15% of the population cholesterol stones (cholelithiasis) are formed.
As noted above, the synthesis of bile acids from cholesterol is regulated by a negative feedback mechanism.
Bile is the main way of removing bilirubin from the body. Bilirubin - The final product of the decomposition of heme, in its chemical structure is tetrapyrrole. The largest amount of bilirubin (80-85%) comes from the decaying hemoglobin of the aging red blood cells and the prematurely collapsing newly formed red blood cells in the bone marrow or circulatory bed (the so-called ineffective erythropoiesis). The rest of the bilirubin is formed as a result of the destruction of other hem-containing proteins (for example, cytochrome P-450, etc.) in the liver and much less in the extrahepatic tissue. It should be noted that the erythrocyte hemoglobin catabolism into bilirubin primarily occurs in the macrophages of the spleen, liver and bone marrow.
Sources of bilirubin formation (in percentage)

As is known, unconjugated bilirubin is hydrophobic (insoluble in water) and a potentially toxic substance that circulates in the plasma in a state closely associated with albumin and cannot be excreted in the urine. The body's ability to eliminate bilirubin is associated with the removal of the latter from blood plasma by the liver cell, subsequent conjugation with glucuronic acid and the release into the bile of an already water-soluble compound (bound or direct bilirubin). The conjugation process proceeds via the microsomal enzyme uridine diphosphate glucuronyl transferase. Conjugation of bilirubin with glucuronic acid has the most important physiological significance; however, conjugation of bilirubin with sulfates, glucose and xylose occurs in small quantities.
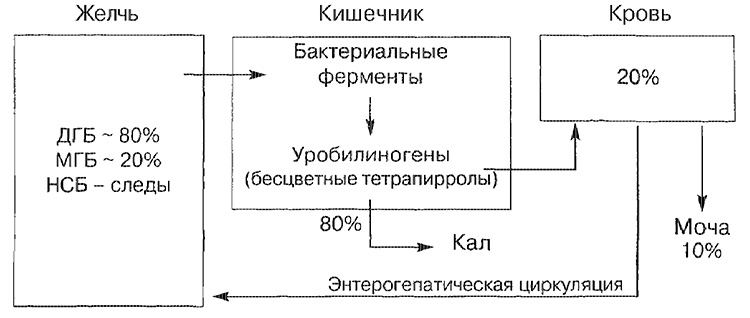
Main transport routes for bilirubin
NSB - unbound bilirubin; MGB - bilirubin monoglucuronide; DGB - diglucuronide bilirubin.
The secretion of conjugated bilirubin into the bile tubules occurs with the participation of a family of ATP-dependent multispecific transport proteins for organic anions.
Most bilirubin bilirubin (about 80%) is represented in the form of diglucuronid bilirubin, a smaller part - in the form of a monoglucuronide, and only a small amount is represented by the unbound form. Bilirubin, which entered the intestine, is broken down in the terminal part of the small intestine and in the large intestine by bacterial enzymes (beta-glucuronidase), which is converted into colorless tetrapyrrole (urobilinogen). About 20% of the formed urobilinogens are resorbed and necessarily excreted in the urine, as well as in the bile (enterohepatic circle of circulation).
Most authors agree that the normal serum total bilirubin concentration determined by the Van Der Bergh diazo reaction usually does not exceed 1 mg% (0.3–1 mg%, or 5–17 µmol / l). Only less than 5% of bilirubin is presented in a bound form. An increase in the level of bilirubin in the blood (hyperbilirubinemia) and its accumulation in the tissues lead to the appearance of jaundice, which, as a rule, becomes noticeable at values in excess of 2.5–3 mg%.
As noted above, bilirubin circulates in the blood in a protein-bound state and hardly penetrates tissue fluids with a low protein content. In this regard, exudates are more yellow in color than transudates. Bilirubin binds well to elastic tissue, which explains the early yellow staining of the sclera, skin and vascular walls in hyperbilirubinemia. Prolonged cholestasis leads to the appearance of a greenish skin tone, which is explained by the deposition of biliverdin.
It will be appropriate to emphasize that the pressure in the bile ducts, against which bile secretion occurs, normally amounts to 150-200 mm of water. Art. Its increase to 350 mm of water. Art. inhibits the secretion of bile, leading to the development of jaundice. In the case of complete cessation of secretion of bilirubin and bile acids, bile becomes discolored (the so-called white bile).
In addition, bile is a way of excretion vegetable fats, lipophilic drugs and their metabolites, various xenobiotics that are present in plants, lipophilic metabolites, fat-soluble vitamins and steroid hormones.
Iron and copper balance the body is also supported due to the biliary excretion of these metals. Both cations are secreted into the bile by an ATP-stimulated canalicular pump. With the bile occurs and the release of other metals.
Bile excretion
The current of bile in the biliary system is caused by the difference in pressure in its various sections and duodenum, sphincter tone, contraction of the smooth muscle fibers of the gastrointestinal tract and ducts (the first individual muscle fibers appear in the intrahepatic bile ducts located in the immediate vicinity of the gates of the liver). The activity of the entire system is normally well coordinated and under the control of the nervous and humoral mechanisms.
In gallstones, bile is deposited and increased in concentration with a 10-fold increase in the concentration of lipids, bile salts, bile pigments, etc. At the right moment, it is released into the OZhP and further into the duodenum. The passage of bile coordinates sphincter apparatus. Outside the digestive process in the duodenum, the sphincter of the vial is closed. At this time, the RI and the cystic duct are relaxed, which allows the bile produced by the liver to enter the lumen of the RI. The mucous membrane ZH absorbs water, ions. At the same time bile becomes more concentrated. Products of mucus allows bile to be in a colloidal state.
The pressure in the lumen of the gland at rest is much less than in the bile ducts, and is 60-185 mm of water. Art. The difference in pressure is the physiological basis of the flow of bile in the gallbladder with the closed Oddi sphincter. In the process of digestion due to the reduction of GF pressure increases to 150-260 mm of water. st, ensuring the flow of bile into the duodenum through the relaxed sphincter ampoules. When bile begins to flow into the duodenum, the pressure in the ducts gradually decreases (with each meal, the fat content decreases 1-2 times).
The period of the primary reaction of the biliary system, due to the type, smell of food and its reception, lasts about 7-10 minutes. Then comes the evacuation period (the main or the period of emptying ZHP), during which, against the background of the alternation of contractions and relaxations of the ZH, the gallbladder and then the hepatic bile flow out into the duodenum.
Reflex stimulation of cholekinesis (conditionally and unconditionally-reflex) occurs due to receptors located in the oral cavity, the stomach and duodenum, through the vagus nerves. Humoral regulation is mainly carried out under the influence of cholecystokinin, which has a stimulating effect on the GF, causing its reduction. In addition, weak cuts in GI occur under the influence of gastrin, secretin, and Bombezin. On the contrary, the contraction of GH, glucagon, calcitonin, anticholecystokinin, vasointestinal peptide and pancreatic polypeptide inhibit.
Egg yolks, fats, milk and meat are powerful stimulants of bile excretion.
Disorders of the sphincter apparatus and gonadal motility can lead to changes in the direction and speed of bile flow, dyskinesia, reflux of pancreatic secretions and the duodenum contents in ACP, and bile in the pancreatic duct, which is the basis for the development of a number of pathological conditions and diseases.
Bile salts, conjugated bilirubin, cholesterol, phospholipids, proteins, electrolytes and water are secreted by hepatocytes into the biliary canaliculi. Bile secretion apparatus includes transport proteins tubular membrane, intracellular organellesand structures cytoskeleton. Tight contactsbetween the hepatocytes, the lumen of the tubules is separated from the circulatory system of the liver.
The tubular membrane contains transport proteins for bile acids, bilirubin, cations and anions. Microvilli increase its area. Organelles are represented by the Golgi apparatus and lysosomes. With the help of vesicles, proteins (eg, IgA) are transported from the sinusoidal membrane to the canalicular membrane, the transport of transport proteins synthesized in the cell for cholesterol, phospholipids and, possibly, bile acids from microsomes to the tubular membrane.
The hepatocyte cytoplasm around the tubule contains the structure of the cytoskeleton: microtubules, microfilamentsand intermediate filaments.
Microtubules are formed by polymerization of tubulin and form a network inside the cell, especially near the basolateral membrane and the Golgi apparatus, taking part in receptor-mediated vesicular transport, secretion of lipids, and under certain conditions - bile acids. Microtubule formation is inhibited by colchicine.
Interacting polymerized (F) and free (G) actins participate in the construction of microfilaments. The microfilaments concentrating around a canalicular membrane determine contractility and motility of tubules. Phalloidin, which enhances actin polymerization, and cytochalasin B, which weakens it, inhibit the motility of the tubules and cause cholestasis.
Intermediate filaments consist of cytokeratin and form a network between the plasma membranes, nucleus, intracellular organelles and other structures of the cytoskeleton. The rupture of intermediate filaments leads to disruption of intracellular transport processes and obliteration of the lumen of the tubules.
Water and electrolytes affect the composition of the tubular secretion, penetrating through tight contacts between the hepatocytes due to the osmotic gradient between the lumen of the tubules and Disse spaces (paracellular current).The integrity of tight contacts depends on the presence on the inner surface of the plasma membrane of the ZO-1 protein with a molecular mass of 225 kDa. The rupture of tight contacts is accompanied by the release of dissolved larger molecules into the canaliculi, which leads to a loss of osmotic gradient and the development of cholestasis. At the same time regurgitation of canalicular bile in sinusoid can be observed.
The biliary canaliculi flow into the ductules, sometimes called cholangiols or the Goering channels. Ductulas are located mainly in the portal zones and flow into the interlobular bile ducts, which are the first of the biliary tracts accompanied by branches of the hepatic artery and portal vein and are found in the composition of the portal triads. The interlobular ducts, merging, form the septal ducts until two main hepatic ducts form, emerging from the right and left lobes in the portal fissure of the liver.
Bile secretion
The formation of bile occurs with the participation of a number of volatile transport processes. Its secretion is relatively independent of perfusion pressure. The total current of bile in humans is approximately 600 ml / day. Hepatocytes provide secretion of two fractions of bile: dependent on bile acids ("225 ml / day) and not dependent on them (" 225 ml / day). The remaining 150 ml / day are secreted by the bile duct cells.
The secretion of bile salts is the most important factor in the formation of bile. (fraction dependent on bile acids).Water moves in the wake of osmotically active salts of bile acids. Changes in osmotic activity can regulate the flow of water into the bile. There is a clear correlation between the secretion of salts of bile acids and the flow of bile.
The existence of the bile fraction, which does not depend on bile acids, is proved by the possibility of the formation of bile that does not contain salts of bile acids. Thus, it is possible to continue the flow of bile, despite the absence of excretion of salts of bile acids; the secretion of water in this case is due to other osmotically active soluble substances, such as glutathione and bicarbonates.
Cellular mechanisms of bile secretion
A hepatocyte is a polar secretory epithelial cell having a basolateral (sinusoidal and lateral) and apical (tubular) membranes.
The formation of bile includes the capture of bile acids and other organic and inorganic ions, their transport through the basolateral (sinusoidal) membrane, cytoplasm and tubular membrane. This process is accompanied by osmotic filtration of water contained in the hepatocyte and paracellular space. The identification and characterization of the transport proteins of the sinusoidal and tubular membranes are complex. Especially difficult is the study of the secretory apparatus of the tubules, however, to date, a technique for obtaining doubled hepatocytes in a short-lived culture has been developed and proven reliable in many studies .. The cloning of transport proteins allows us to characterize the function of each of them separately.
The process of bile formation depends on "the presence of certain carrier proteins in the basolateral and tubular membranes. Na +, K plays the role of the driving force of secretion + - ATPase of the basolateral membrane, providing a chemical gradient and potential difference between the hepatocyte and the surrounding space. Na +, K + - ATPase exchanges three intracellular sodium ions for two extracellular potassium ions, maintaining a concentration gradient of sodium (high outside, low inside) and potassium (low outside, high inside). As a result, the cell content has a negative charge (–35 mV) compared with the extracellular space, which facilitates the capture of positively charged ions and the excretion of negatively charged ions. Na +, K + -ATPase is not detected in the tubular membrane. Membrane fluidity can affect enzyme activity.
Capture on the surface of a sinusoidal membrane
The basolateral (sinusoidal) membrane has many transport systems for capturing organic anions, the substrate specificity of which partially overlaps. The characterization of carrier proteins was previously given on the basis of studying animal cells. The recent cloning of human transport proteins has made it possible to better characterize their function. The transport protein for organic anions (organic anion transporting protein - OATP) is sodium-independent, carries molecules of a number of compounds, including bile acids, bromsulfalein and, probably, bilirubin. It is believed that the transport of bilirubin in the hepatocyte is also carried out by other carriers. Capture of bile acids conjugated with taurine (or glycine) is carried out by sodium / taurocholate transport protein (sodium / bile acid cotransporting protein - NTCP).
Ion transfer through the basolateral membrane involves a protein that exchanges Na + / H + and regulates the pH inside the cell. This function also performs cotransport protein for Na + / HCO 3 -. On the surface of the basolateral membrane, the capture of sulphates, non-esterified fatty acids, and organic cations also occurs.
Intracellular transport
Transport of bile acids in the hepatocyte is carried out using cytosolic proteins, among which the main role is played by Z-hydroxysteroid dehydrogenase. Glutathione-S-transferase and fatty acid binding proteins are of lesser importance. The endoplasmic reticulum and the Golgi apparatus are involved in the transfer of bile acids. Apparently, vesicular transport is activated only when bile acids are significantly injected into the cell (at concentrations exceeding physiological).
The transport of proteins in the liquid phase and ligands, such as IgA and low density lipoproteins, is carried out by vesicular transcytosis. The transfer time from the basolateral to the tubular membrane is about 10 minutes. This mechanism is responsible only for a small part of the total current of bile and depends on the state of the microtubules.
Tubular secretion
The tubular membrane is a specialized portion of the hepatocyte plasma membrane containing transport proteins (mostly ATP-dependent) responsible for the transfer of molecules into bile against a concentration gradient. Enzymes such as alkaline phosphatase, GGTP are also localized in the canalicular membrane. The transfer of glucuronides and glutathione-S-conjugates (for example, bilirubin diglucuronide) is carried out with the help of a canalicular multispecific transport protein for organic anions (cMOAT), the transport of bile acids with a canalic transport protein for bile acids (canalicular b transporter - SWAT), whose function is partially controlled by negative intracellular potential. The current of bile, which does not depend on bile acids, is apparently determined by the transport of glutaion, as well as the canalicular secretion of bicarbonate, possibly with the participation of a protein exchanging Cl - / HCO 3 -.
An important role in the transport of substances through the tubular membrane belongs to two enzymes of the P-glycoprotein family; both enzymes are ATP-dependent. Multidrug resistance protein 1 (MDR1) multidrug resistance protein 1 transfers organic cations and also removes cytostatic drugs from cancer cells, causing their resistance to chemotherapy (hence the name protein). The endogenous substrate of MDR1 is unknown. MDR3 transports phospholipids and acts as a flip-up for phosphatidylcholine. The function of MDR3 and its importance for the secretion of phospholipids in bile are refined in experiments on mice that lack the mdr2-P-glycoprotein (analogous to human MDR3). In the absence of phospholipids in the bile, bile acids cause damage to the biliary epithelium, inflammation of the ductules and periductular fibrosis.
Water and inorganic ions (especially sodium) are excreted into the biliary capillaries along the osmotic gradient by diffusion through negatively charged semipermeable tight contacts.
Bile secretion is regulated by many hormones and secondary messengers, including cAMP and protein kinase C. Increasing the concentration of intracellular calcium inhibits bile secretion. The passage of bile through the tubules is due to microfilaments, which provide motility and contraction of the tubules.
Ductile secretion
Epithelial cells of the distal ducts produce a bicarbonate-rich secret that modifies the composition of the tubular bile (the so-called ductular current, bile).The secretion process produces cAMP, some membrane transport proteins, including a protein that exchanges Cl - / HCO 3 -, and regulator of transmembrane conduction in cystic fibrosis -membrane channel for Cl -, adjustable cAMP. Ductular secretion is stimulated by secretin.
It is assumed that ursodeoxycholic acid is actively absorbed by the ductile cells, exchanged for bicarbonates, recycled in the liver and subsequently excreted again into the bile ("cholehepatic shunt"). Perhaps this explains the choleretic effect of ursodeoxycholic acid, accompanied by a high biliary bicarbonate secretion in experimental cirrhosis.
The pressure in the bile ducts, at which bile secretion occurs, is normally 15-25 cm of water. Art. Increase pressure to 35 cm water. Art. leads to suppression of bile secretion, development of jaundice. Secretion of bilirubin and bile acids can be completely stopped, while the bile becomes colorless (white gall)and resembles mucous fluid.
Bile is an isosmotic plasma liquid consisting of water, electrolytes and organic substances (bile acids, phospholipids, cholesterol, bilirubin). Bile acids (or their salts) are the main organic component of bile. Bile acids enter bile from two sources: (1) primary bile acids (cholic and chenodeoxycholic), which are synthesized from cholesterol in the liver; (2) secondary bile acids (deoxycholic, lithocholic and ursodeoxycholic) are formed by the action of intestinal bacteria from primary bile acids. The bile acids themselves consist of two important components, which determine their physiological and physicochemical properties: (1) a steroid core with hydroxyl substituents; (2) an aliphatic side chain (Figure 7-4).
Fig. 7-4.
Bile acids consist of two components - a nucleus with hydroxyl termini and an aliphatic side chain. In the figure, cholic acid is shown as an example of a three-hydroxy acid (3? -, 7? -, 12? -OH). Other examples are bile acids containing deoxycholate (3? -, 12? -OH), chenodeoxycholate (3? -, 7? -OH) and lithocholate (3? -OH)
In most mammals, primary bile acids contain from three to seven hydroxyl substituents, the number of which affects their water solubility (hydrophilicity). Soon after the formation, the primary bile acids undergo modification in the terminal carboxyl group. This occurs during the hepatic phase of the enterohepatic circulation of secondary bile acids and their conjugation with glycine or taurine. The presence of hydrophilic (hydroxyl components and amide bonds of the aliphatic side chain) and hydrophobic (steroid core) components allows conjugated bile acid molecules to act as an amphoteric compound. This gives them the opportunity to form micelles (polymolecular aggregates) above the critical micellar concentration. In turn, molecules of bile acids are able to dissolve other amphoteric substances (cholesterol, phospholipids) with the formation of mixed micelles. This detergent-like role of bile acids is important for stabilization of the physico-chemical state of bile, digestion and absorption of fats.
Synthesis of bile acids from cholesterol is regulated by a negative feedback mechanism, although the nature of regulation at the molecular and biochemical levels is not yet fully understood. Microsomal 7? -Hydroxylation of cholesterol is a critical step in the synthesis of bile acids. Chenodesoxycholic acid, used to dissolve gallbladder stones, inhibits the synthesis of bile acids and thereby increases blood cholesterol levels. When using ursodeoxycholic acid, such changes are not observed even with long-term treatment.
The formation of bile takes place both on the sinusoidal and tubular surface of the hepatocyte membrane and is both an intracellular and paracellular process. Unlike glomerular filtration in the kidneys, which passively under the action of hydrostatic forces, during the formation of bile, there is an active transfer of organic and inorganic components into the lumen of the tubules and passive transport of water. Thus, the processes of bile secretion are similar to the processes of secretion in the acini of the pancreas, the epithelium of the renal tubules. The formation of tubular bile can be divided into two types (Fig. 7-5): (1) bile formation, dependent on the secretion of bile acids, defined as the ratio of the amount of bile secreted into the tubules to the amount of secreted bile salts; (2) bile formation, independent of bile acid secretion, which can be represented as an active secretion of inorganic electrolytes and other substances and is reflected on the graph as a y-intersection of this line. In other words, bile formation associated with acid secretion is the flow rate of bile, depending on the presence of osmotically active bile salts in the bile ducts, and bile formation unrelated to acid secretion in the absence of bile salts. The ratio of the rate of formation of bile and the formation of salts of bile acids is non-linear in nature with small amounts of secreted bile and cannot correspond to the linear relationship shown in Fig. 7-5. Therefore, both types of bile formation should be considered as interrelated indicators of bile formation.
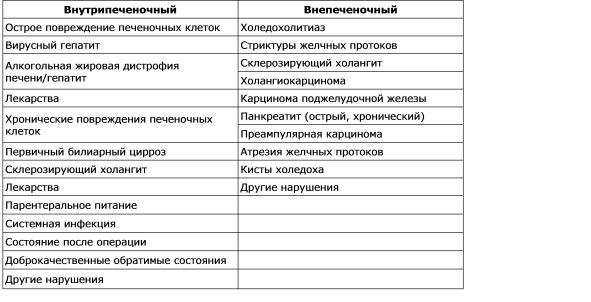
Violation of bile formation is called cholestasis. The sequence of emerging pathological, physiological and clinical manifestations of cholestasis depends on the cause of it. During the morphological study of the material of hepatobiopsy bile is detected in the tubules of pericentral hepatocytes, the dilatation of the tubules is noted, and the study of the ultrastructure reveals a decrease in the number of microvilli. Cholestasis can be defined as a functional defect in the formation of bile at the level of the hepatocyte (intrahepatic cholestasis), as well as organic or mechanical impaired secretion and outflow of bile (extrahepatic cholestasis). The most common causes of intrahepatic and extrahepatic cholestasis are given in Table. 7-2. There are several mechanisms that play an important role in the pathogenesis of intrahepatic cholestasis: impairment and damage to the function of the sinusoidal membrane; violation of the functions of hepatocyte intracellular organelles; damage and disruption of the tubular membrane. Thus, there is no single mechanism for cholestasis in various clinical situations, and the multiplicity of mechanisms can lead to various disorders. Clinically, cholestasis is characterized by an increase in blood levels of many substances, including bilirubin, bile salts, cholesterol, which are normally secreted into bile. In the biochemical analysis of blood with cholestasis, a disproportionate increase in alkaline phosphatase activity and bilirubin concentration is observed in parallel with changes in the level of aminotransferases, which will be discussed below.
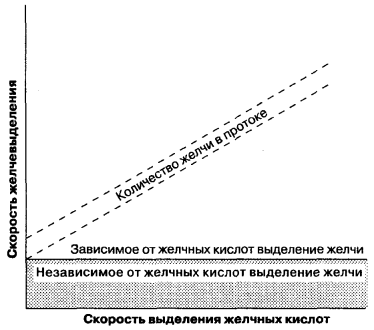
Fig. 7-5.
Bile formation associated with bile acid secretion, bile formation independent of bile acid secretion. (By: Moseley R. H., Bile secretion. In: Yamada T., Alpers D. H., Owyang C., Powell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995: 387.)
Table 7 - 2.
In the liver, the most important digestive juice is formed - bile.
Bile is produced by hepatocytes by active and passive transport of water, cholesterol, bilirubin, cations into them. In hepatocytes from cholesterol, primary bile acids are formed - cholic and deoxycholic. A water-soluble complex is synthesized from bilirubin and glucuronic acid. They enter the bile capillaries and ducts, where bile acids combine with glycine and taurine. As a result, glycocholic and taurocholic acids are formed. Sodium bicarbonate is formed by the same mechanisms as in the pancreas.
Bile is produced by the liver all the time. In its day about 1 liter is formed. Hepatocytes excrete primary or hepatic bile. This liquid is a golden yellow alkaline reaction. Its pH is 7.4 - 8.6. It consists of 97.5% water and 2.5% solids. The dry residue contains:
1. Mineral substances. Cations of sodium, potassium, calcium, bicarbonate, phosphate anions, chlorine anions.
2. Acidic acids - taurocholic and glycocholic.
3. Bile pigments - bilirubin and its oxidized form biliverdin. Bilirubin gives bile color.
4. Cholesterol and fatty acids.
5. Urea, uric acid, creatinine.
Since outside the digestive system, the sphincter of Oddi, located at the mouth of the common bile duct, is closed, the excreted bile accumulates in gallbladder. Here water is reabsorbed from it, and the content of basic organic components and mucin increases by 5-10 times. Therefore, cystic bile contains 92% water and 8% dry residue. It is darker, thicker and more viscous than the liver. Due to this concentration, the bladder can accumulate bile for 12 hours. During digestion, the sphincter of Oddi and the sphincter of Lutkens in the bladder neck open. Bile enters the duodenum.
Bile Value:
1. Malic acids emulsify a part of fats, turning large fat particles into fine droplets.
2. It activates the enzymes of intestinal and pancreatic juice, especially lipase.
3. In combination with bile acids, long-chain fatty acids and fat-soluble vitamins are absorbed through the enterocyte membrane.
4. Zhelchka promotes resynthesis of triglycerides in enterocytes.
5.Inactivates pepsins, and also neutralizes the sour chyme coming from the stomach. This ensures the transition from gastric to intestinal digestion.
6.Stimulates the secretion of pancreatic and intestinal juices, as well as the proliferation and desquamation of enterocytes.
7. Strengthens intestinal motility.
8. It has a bacteriostatic effect on intestinal microorganisms and thus prevents the development of putrefactive processes in it.
The regulation of bile formation and biliary excretion is mainly carried out by humoral mechanisms, although nervous ones play a certain role. The most powerful stimulator of bile formation in the liver is bile acids absorbed into the blood from the intestine. It is also enhanced by secretin, which contributes to an increase in sodium bicarbonate in bile. The vagus nerve stimulates the production of bile, the sympathetic inhibit.
When the chyme enters the duodenum, I-cells begin to release its cholecystokinin-pancreozymin i-cells. Especially this process is stimulated by fats, egg yolk and magnesium sulfate. KTK-PZ strengthens contractions of smooth muscles of the bladder, bile ducts, but relaxes the sphincters of Lutkens and Oddi. Bile is released into the gut. Reflex mechanisms play a small role. Chyme irritates chemoreceptors of the small intestine. Impulses from them enter the digestive center of the medulla oblongata. From him they are on the vagus to the biliary tract. The sphincters relax, and the smooth muscles of the bladder contract. It promotes biliary excretion.
The most serious diseases are hepatitis and cirrhosis. Most often, hepatitis is the result of infection (infectious hepatitis A, B, C) and exposure to toxic products (alcohol). In hepatitis, hepatocytes are affected and all liver functions are impaired. Cirrhosis is the outcome of hepatitis. The most common violation of biliary excretion is cholelithiasis. The bulk of gallstones are formed by cholesterol, since the bile of such patients is supersaturated with them.
The liver is the largest organ of the body, which is central to the metabolism. It performs many functions, participating in the metabolism of proteins, carbohydrates, fats, hormones and vitamins, as well as in the neutralization of many endogenous and exogenous substances. These processes are described in physiological chemistry textbooks, and in this section, in connection with digestive system we will consider only excretory liver functionthose. bile secretion. Bile consists of water, mineral salts, mucus, cholesterol lipids and lecithin and two types of specific components - bile acids and bilirubin pigment. Bile acids are detergents, and their emulsifying action plays an important role in lipid digestion. Bilirubin is the end product of the breakdown of hemoglobin to be eliminated from the body.
^
Bile formation
Functional anatomy.Liver cells (hepatocytes)form a plate in the thickness of one cell, separated by narrow slits (Disse space)representing the completed
^ 766 PART VIII. FOOD, DIGESTION AND EXTRACT
By blood sinusoidsequivalent to capillaries. In the walls of sinusoids there are pores through which such large macromolecules as albumin and lipoproteins can pass. In the smallest tubulesbile capillaries bounded by the plasma membranes of two adjacent hepatocytes - bile is collected in larger goring's tubules,the walls of which, as well as larger in size intercollegeular tubulesand bile ductsformed by cubic secretory cells. The small tubules inside the liver lobules and between them merge into larger ones, eventually forming hepatic duct.From this duct departs cystic ductto the gallbladder. After the merger, the hepatic and cystic ducts form common bile ductopening into the duodenum at the top of the Vater papilla behind or near the pancreatic duct (Fig. 29.1).
^ Functions of bile.Bile performs many important functions. Together with her end products are displayedexchange, such as bilirubin, as well as drugs and toxins. Allotment withbile cholesterolplays an important role in the regulation of its balance. Bile acidsessential for emulsification and fat absorption. In addition, bile contains water, mineral salts and mucus. About 24 hours a day 600 mlbile, and 2/3 of this amount comes from the tubules, and 1/3 of the larger ducts.
^ Tubular bile formed in approximately equal amounts with the participation of two different mechanisms, bile-dependent and bile-dependent (fig. 29.27).
^ Bile acid dependent secretion. There is a close relationship between bile excretion rateand bile acid secretion.In canalicular bile, the concentration of bile acids is 100 times higher than in portal blood; therefore, they are considered to be secreted by active transport involving vector.Following the bile acids along the osmotic gradient, water rushes into the tubules, so the bile is isotonic with blood.
There are two sources of bile acids. First, they are synthesized de novo from cholesterol in the hepatocytes themselves with the participation of 7-hydroxylase. This enzyme plays a key role and controls the rate of synthesis of bile acids through a feedback mechanism. Secondly, hepatocytes are able to actively absorb bile acids from portal bloodand excrete them into the tubules (see also fig. 29.29). This extraction is very effective; with a single passage of blood through the liver, 80% of bile acids are extracted from it. therefore
The concentration of bile acids in peripheral blood is much lower than in the portal system. Since bile acids are extracted from the blood 6 times faster than they enter the tubules, it is the latter process that limits the rate of bile acid secretion.
^ Secretion independent of bile acids. AT
This process involves ions Na +, Cl -, HCO 3 - and water. The driving force is active transport Να + , possibly with bicarbonate. A secretion independent of bile acids stimulates, in particular, secretin.
In addition to bile acids, bilirubin, cholesterol and phospholipids (mainly lecithin) are actively secreted into the canaliculi (Fig. 29.27). Water insoluble ("Indirect") bilirubin,most of which is formed from hemoglobin of aged red blood cells, enters the hepatocytes in the form of a colloidal aggregate associated with albumin. Its daily formation is about 4 g / kg body weight, or 200-300 mg / day. In hepatocytes, 80% of bilirubin is conjugated. withglucuronic acid and a small amount of it with sulfuric acid. In such conjugated
^ CHAPTER 29. FUNCTIONS OF THE GASTROINTESTINAL TRACT 767
Bilirubin is excreted in the bile. ("Direct" bilirubin).Basically, drugs and toxins are removed in the same way.
^ Modification of bile in the bile ducts
(fig. 29.27). In the ducts, where the tubules open, the primary bile undergoes modification. This process resembles the process of modifying the glomerular filtrate in the renal tubules (p. 785), and in a similar way is calculated for it clearance,only in the case of bile as an inert substance is used instead of inulin erythritolor mannitol,which are secreted into the tubules, but not reabsorbed. Similar studies have shown that about 180 ml of bile, or 1/3 of its total amount, is secreted into the ducts during active secretion of NSO ^ ". This process is stimulated secretin
^
Hepatic and gallbladder bile
The composition of hepatic bile(tab. 29.3). The bile secreted by the liver at a rate of 0.4 ml / min has a golden color, which is explained by the presence of bilirubin in it. The concentration of electrolytes in this bile is the same as in plasma, except for the fact that it contains twice as much HCO3 "and somewhat less than CI ~. At the same time, the composition of organic substances bile is very different from plasma, since in bile they are represented almost exclusively by bile acids, cholesterol and phospholipids.
 |
^ Bile acidsare formed in the liver from cholesterol as a result of its hydroxylation and the addition of a carboxyl group. Acids in the liver are primary bile acids;these include chenodeoxycholic(dioxyform) and holevaya(trioxyforma) acid.In the liver, they are not in free form, but in the form of conjugates with glycine and taurine, and conjugates with glycine are formed three times more, since the amount of taurine is limited. Conjugated bile acids are more soluble in water,
Than unconjugated, and have a greater ability to dissociate and form bile salts with cations, mainly with Na + ions. In an acidic medium (pH 4.0), bile acid salts are insoluble and precipitate, but at physiological pH values (in the small intestine), they are well soluble.
In the distal part of the ileum and in the colon, some of the salts of the primary bile acids undergo dehydroxylation under the action of anaerobic bacteria and turn into secondary bile acid-lithocholic(monooxyform) and deoxycholic(dioxyform). Chenodeoxycholic, cholic and deoxycholic acids are present in a 2: 2: 1 ratio. Litocholic acid is present only in some fractions, since most of it is excreted.
The emulsifying effect of bile acids on fats is based mainly on their ability to form micelles.The bile acid molecules have a three-dimensional structure in which the hydrophilic carboxyl and hydroxyl groups are on the same side of the molecule, and the hydrophobic part of the molecule (steroid core, methyl groups) is on the opposite side, due to which the bile acid molecules have and hydrophilicand lipophilic properties.Due to this structure, the molecules of bile acids act as detergents: at the interface of the lipid and aqueous phases, they form an almost monomolecular film in which the hydrophilic groups are turned to the aqueous, and the lipophilic to the lipid phase. In the aqueous phase, bile acids form ordered aggregates. -micelsprovided that their concentration reaches a certain level, called critical micelle concentration(1-2 mmol / l). The inner, lipophilic region of the micelle may contain lipids,for example, cholesterol and phospholipids; such micelles are called mixed (Fig. 29.28). By itself, cholesterol is insoluble in water, but in the composition of micelles can be in solution. If its concentration exceeds the capacity of micelles, it forms a crystalline precipitate; this process underlies the formation of cholesterol gallstones (p. 769).
^ Composition of cystic bile (tab. 29.3). The capacity of the gallbladder is only 50-60 ml. At the same time, the liver secretes bile at a rate of 600 ml / day, and half of this amount passes through the gall bladder before entering the small intestine. The difference between the volume of bile entering the gallbladder and its capacity is compensated highly efficient reabsorptionwater in the gallbladder. Within a few hours, 90% of the water can be reabsorbed from the bile. With
^
768 PART VIII. FOOD, DIGESTION AND EXTRACT
This organic matter remains in the gallbladder and their concentration in the bile increases. The driving force of reabsorption is active transport of Nα ions *with the participation of a “pump” embedded in the basal and lateral membranes of cells and activated by (Na + -K +) -ATPhase. Following Na + ions, Cl - and HCO 3 - ions diffuse in the direction of the electric gradient or transported by carriers move. As a result of the reabsorption of HCO 3, the pH of the gallbladder decreases to 6.5 vs 8.2 in hepatic bile. Due to the creation in the extracellular space of the epithelium of the gallbladder a high concentration of Na + ions, an osmotic gradient occurs, leading to the pumping of water, which then flows into the capillaries (p. 751).
^ Motility gallbladder. In the fasting state, bile accumulates in the gallbladder, and in
Eating time is allocated as a result of contractions of the gallbladder. The main stimulator of the contractile activity of the gallbladder is cholecystokinin,secreted by the mucous membrane of the duodenum upon admission to it of chyme containing fat. To some extent, contractions of the gallbladder are also stimulated. vagus nerveand parasympatholytics.They begin within 2 minutes after fatty food comes in contact with the intestinal mucosa, and after 15-90 minutes the bladder is completely emptied. Motility of the gallbladder involves two processes. Initially, a tonic contraction develops, as a result of which the diameter of the gallbladder decreases, and then periodic contractions are imposed on this effect, the frequency of which is 2-6 / min. As a result of these two processes, a pressure of 25–30 mm Hg is created. Art.
^
Enterohepatic circulation
Bile acid circulation(fig. 29.29). Bile acids are secreted into the duodenum as mixed micelles. Despite the dilution of bile acids by the contents of the stomach, their concentration in the intestine is about 10 mmol / l and remains above the critical concentration of micelle formation. Here, in addition to cholesterol and lecithin, micelles include the products of hydrolytic fat breakdown - fatty acidand monoglycerides.Upon initial contact of the micelles with the intestinal wall
^
CHAPTER 29. FUNCTIONS OF THE GASTROINTESTINAL TRACT 769
Lipids diffuse through the membrane of the brush border into enterocytes, and bile acids remain in the intestinal lumen, but with further passage through the intestine, bile acids are absorbed by active and passive transport.
About 50% of bile acids are absorbed in the intestine passively. As a result of the cleavage of bile acid conjugates and the dehydroxylation of the latter by the action of intestinal bacteria, their solubility in lipids increases and passive diffusion is facilitated.
^ Active suction bile acids occur exclusively in the terminal ileum - a rare phenomenon, known only for the absorption of vitamin B 12. Only those bile acids that have a high polarity that impede their passive absorption, such as taurine conjugates, are subject to active absorption. The process of absorption of bile acids in the terminal ileum is characterized by typical signs of active transport: saturation kinetics and competitive inhibition. A small amount of bile acids (7-20%) is not included in either active or passive absorption and is eliminated from the body.
The presence of bile acids in the colon plays an important role in regulating the consistency of feces. When the concentration of Dioxy Acid in the colon over 3 mmol / l, a significant amount of electrolytes and water is secreted into the intestinal lumen, which leads to diarrhea. Pronounced form of this "Hologennogo" diarrheacan be observed with resection or disease of the terminal ileum, and for its treatment using the binding of bile acids using cholestyramine ion exchanger.
When ingested absorbed bile acids to the liverconjugates are newly formed, and some secondary bile acids undergo hydroxylation. Loss of bile acids in the feces (0.2-0.6 g / day) is compensated by their synthesis.
^ Total pool of bile acids in the body is about 3.0 g. This amount is not enough to ensure lipolysis after meals; in particular, the consumption of fatty foods requires 5 times more bile acids. However, the body is not deficient in bile acids, because they circulate through the intestines and the liver many times. (enterohepatic circulation).The frequency with which the pool of bile acids makes a full cycle depends on the dietary regime and ranges from 4 to 12 cycles per day.
^ Circulation of bilirubin. Gall pigment bilirubin,like bile acids and lipids, enters the intestine in the form of glzhuronid. Only a small amount of this polar compound
Reabsorbed in the gallbladder and small intestine. In the terminal ileum and (mostly) in the colon, the bilirubin conjugate is cleaved by bacterial hydrolases. At the same time bilirubin turns into urobilinogenwhich, along with other decay products of bilirubin, gives brown color to feces. Less than 20% of urobilinogen is absorbed back, and of this amount, about 90% re-enters the liver and returns to bile, and the remaining 10% is excreted in the urine.
Pathophysiological aspects. Elevated levels of urobilinogen in the urine may indicate liver disease,accompanied by a violation of bilirubin excretion. The complete absence of urobilinogen in the urine, the light color of feces and yellowness indicate complete blockage of the gallbladder;in this situation, bilirubin does not enter the intestine at all and urobilinogen is not formed.
The most well-known and widespread violation of the normal physiology of the biliary system is the precipitation of cholesterol with the formation of cholesterol gallstones. Cholesterol, like lecithin, is in a dissolved state only as part of mixed micelles. If the concentration increases xo.jecmepo.ia or concentration of bile acids or lecithin becomes below critical levels,cholesterol precipitates. Among the factors causing an increase in the relative cholesterol content are estrogens, a carbohydrate diet, overweight, and processes that reduce the concentration of bile acids, such as inflammation of the ileum. (Crohn's disease)or resection. In some cases, oral bile acid supplementation is enough to turn lithogenic bile into alitogenic, in which cholesterol stones can dissolve. Chenodeoxycholic and urodezoxycholic acids are most suitable for this purpose, since they do not cause diarrhea.
The clinical manifestation of impaired bilirubin metabolism is jaundice. Yellow skin is associated with elevated plasma levels of bilirubin, which can occur in the following cases:
with increased formation of bilirubin as a result of enhanced erythrocyte breakdown (hemolytic jaundice);
as a result of disruption of the conjugation process or transport of bilirubin in hepatocytes, as, for example, jaundice pregnantor gilbert's maternity jaundice;
with delayed outflow of bile, for example due to gallstones or tumors localized in the region of the bile duct (obstructive jaundice).